
Zwangerschap



Zwangerschap (graviditeit) is de toestand waarin een vrouw een bevruchte eicel (zygote), en vervolgens embryo en foetus, in zich draagt.
Meetschalen van de zwangerschapsduur[bewerken | brontekst bewerken]
Om de fase in een zwangerschap uit te drukken zijn twee 'leeftijden' van belang (twee meetschalen):[1]
- de postmenstruele leeftijd (PML): de verstreken tijd sinds de eerste dag van de laatste menstruatie. De zwangerschapsduur wordt meestal uitgedrukt als de postmenstruele leeftijd in weken. Als iemand "tien weken zwanger is", betekent dit dus over het algemeen dat de PML tien weken is. Gemiddeld vindt de bevalling plaats bij een PML van 40 weken. Aan het begin van de zwangerschap wordt op basis daarvan de datum bepaald waarop de bevalling naar schatting plaatsvindt (uitgerekende datum, in statistische termen de verwachtingswaarde van de datum van de bevalling).
- de conceptionele leeftijd (in het Engels fertilization age, ook aangeduid als de 'embryonale leeftijd' en later 'foetale leeftijd'): de leeftijd van een embryo bij een zwangerschap vanaf het moment van de conceptie (bevruchting). De conceptionele leeftijd wordt over het algemeen gebruikt in de beschrijving van de prenatale ontwikkeling van een foetus en in de neonatologie in het algemeen. Het moment van bevruchting wordt over het algemeen geschat op ongeveer twee weken na de eerste dag van de laatste menstruatie. Als iemand volgens de PML "tien weken zwanger is", is de conceptionele leeftijd dus over het algemeen acht weken. Gemiddeld vindt de bevalling plaats bij een conceptionele leeftijd van 38 weken.
Men kan de PML in eerste instantie meestal exacter vaststellen dan de conceptionele leeftijd. In een latere fase van de zwangerschap kan de conceptionele leeftijd beter worden bepaald dan in het begin, op basis van de grootte van de foetus.
Verloop[bewerken | brontekst bewerken]
Een zwangerschap wordt in het algemeen ingedeeld in drie kwart jaren, zwangerschapstrimesters genoemd. Een trimester komt overeen met dertien weken.
Eerste trimester[bewerken | brontekst bewerken]



De ovulatie (eisprong) vindt gemiddeld twee weken na de eerste dag van een menstruatie plaats, als zwangerschap volgt dus bij een PML van twee weken. Binnen een etmaal daarna vindt dan de bevruchting (conceptie) plaats, waarbij een zaadcel een eicel bevrucht, waardoor er een zygote (bevruchte eicel) wordt gevormd. De zaadcel is over het algemeen afkomstig van een ejaculatie (zaadlozing) in de vagina bij vaginale geslachtsgemeenschap, niet meer dan drie etmalen voor de bevruchting. Circa zes dagen na de bevruchting volgt de innesteling, de nidatie van het embryo (de morula) in het endometrium van de baarmoeder. Na een bevruchting blijven daaropvolgende menstruaties uit. Een zwangerschapstest is over het algemeen zinvol vanaf de dag dat de eerstvolgende menstruatie wordt verwacht: de test is, in het geval van zwangerschap, in het algemeen positief vanaf die dag of 1 à 2 dagen later. Eerder is de kans op een fout-negatieve uitslag groot.
Voor de innesteling deelt de zygote zich in kleinere cellen, een klompje totipotente (nog ongedifferentieerde) cellen dat zich excentrisch in de blastocyste bevindt. Circa zes dagen na de bevruchting, dus bij een PML van 3 weken, zijn er 16 tot 32 cellen en volgt de innesteling (nidatie) van het embryo (de morula) in het baarmoederslijmvlies (endometrium).
Gedurende de eerste drie maanden van een zwangerschap, het eerste zwangerschapstrimester, voelt een deel van de aanstaande moeders zich vaak misselijk, en kan de geur van sommige levensmiddelen, bijvoorbeeld van koffie, als zeer onprettig worden ervaren. Het is mogelijk dat deze zwangerschapsmisselijkheid het embryo beschermt tegen stoffen die schade kunnen berokkenen.[bron?]
Tweede trimester[bewerken | brontekst bewerken]
Tijdens de middelste drie maanden van een zwangerschap voelen de meeste aanstaande moeders zich beter.[bron?] Na ongeveer zestien weken kan de zwangere vrouw de foetus voelen bewegen (schoppen).
Derde trimester[bewerken | brontekst bewerken]
Gedurende de laatste drie maanden gaat het kind in de baarmoeder veel energie vragen: het groeit snel en neemt toe in gewicht. Daardoor voelen aanstaande moeders zich sneller moe. Een zwangere vrouw komt gemiddeld ongeveer twaalf kilo aan, sommigen tot 25 kilo, tijdens de zwangerschap. Slechts ongeveer een derde daarvan is het gewicht van het kind zelf.
De zwangerschap eindigt met de geboorte van het kind. De bevalling vindt plaats bij een postmenstruele leeftijd van gemiddeld 40 weken, dus een conceptionele leeftijd van gemiddeld 38 weken. Een "zwangerschapsduur" (PML bij de bevalling) tussen 37 en 42 weken wordt als normaal beschouwd. Dit wordt de "à terme periode" genoemd.
De foetus gaat op het eind van de zwangerschap het stresshormoon cortisol aanmaken. Dit hormoon komt via de placenta in het bloed van de zwangere vrouw. Onder invloed van dit hormoon en de hormonen oxytocine en prostaglandine worden de weeën opgewekt.
Herstel[bewerken | brontekst bewerken]
Na de geboorte van het kind kan het zes weken tot enkele maanden duren voordat de lichamelijke toestand van de vrouw weer enigszins is teruggekeerd tot die van vóór de zwangerschap. Deze periode heet 'ontzwangering' of puerperium. Borstvoeding speelt een positieve rol in dit proces.
Stadia in beeld[bewerken | brontekst bewerken]
- (aantal maanden refereert aan PML)

-
 1 maand
1 maand -
 2 maanden
2 maanden -
 3 maanden
3 maanden -
 4 maanden
4 maanden -
 5 maanden
5 maanden -
 6 maanden
6 maanden -
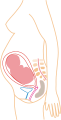 7 maanden
7 maanden -
 8 maanden
8 maanden -
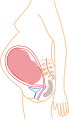 9 maanden
9 maanden









-
 26 en 40 weken
26 en 40 weken -
 27, 29, 31 en 33 weken
27, 29, 31 en 33 weken

Preventie en abortus[bewerken | brontekst bewerken]
Wanneer zwangerschap voorkomen dient te worden en men zich niet wil onthouden van seks, dan kan gebruik worden gemaakt van voorbehoedmiddelen. Bij ongewenste zwangerschap wordt soms abortus toegepast.
Zwangerschap en afweersysteem[bewerken | brontekst bewerken]
Cytokine IL-10 is voor de aanleg van een gezonde zwangerschap en het voorkomen van vroeggeboortes nodig.
B-cellen hebben aan het begin van de zwangerschap een dempend effect op het afweersysteem en vergroten daarmee de tolerantie van het ‘lichaamsvreemde materiaal’ van het embryo dat voor de helft uit lichaamsvreemd materiaal van de partner bestaat.[2]
NK-cellen in de baarmoeder zorgen daar dat het embryo, dat voor de helft 'lichaamsvreemd' niet wordt afgestoten. Ook helpen de NK-cellen mee om de bloedvaten van de moederkoek meer open te zetten, zodat voeding het embryo beter bereikt. Onderzoek liet zien dat de menstruatie van een deel van de vrouwen die miskramen hadden gehad een lager aantal NK-cellen bevatte, vergeleken met vrouwen die geen miskramen hadden gehad.[3]
Verschijnselen bij zwangerschap[bewerken | brontekst bewerken]
Een vrouw in de zwangerschap is kwetsbaar. Bij alle vrouwen doen zich verschijnselen voor die een direct gevolg zijn van de zwangerschap. Dat kunnen zijn:
- Vermoeidheid
- Misselijkheid en braken
- Brandend maagzuur
- Het uitblijven van de menstruatie
- Donker wordende tepels en een donkere, verticale streep op de buik, linea nigra
- Vaak moeten plassen
Innestelingsbloeding[bewerken | brontekst bewerken]
Een aantal zwangere vrouwen krijgt nog een bloeding op het moment dat hun volgende menstruatie had moeten komen. Deze bloeding wordt veroorzaakt door het bevruchte embryo dat zich innestelt in het baarmoederslijmvlies. Zo'n implantatiebloeding gaat doorgaans met minder bloedverlies en minder buikkrampen gepaard. Bloedverlies in het verdere verloop van de zwangerschap kan heel onschuldig zijn, maar men doet er toch goed aan in een dergelijk geval een arts te raadplegen. Het kan immers ook wijzen op een dreigende miskraam, zeker als de bloeding hevig en pijnlijk is.
Problemen, ziekten en syndromen gedurende de zwangerschap[bewerken | brontekst bewerken]
Naar schatting vijftien procent van de vastgestelde zwangerschappen eindigt in een miskraam. Van alle bevruchte eicellen gaat zelfs ongeveer 50% te gronde, maar meestal is dat al vóór de vrouw merkt dat ze zwanger is.
Ziekten en problemen die zich met betrekking tot de zwangerschap kunnen voordoen zijn:
- Bekkenpijn (ook wel bekkeninstabiliteit of symfysiolyse), pijn en instabiliteit in het bekken door weker en rekbaarder worden van verbindingen in het bekken.
- Bloedverlies, veroorzaakt door
- Placentaloslating
- Vasa praevia of voorliggende placenta
- Groeistoornissen, achterstand of voorlopen van de groei.
- Hoge bloeddruk
- Miskraam of een abortus
- Polyhydramnion, te veel vruchtwater of oligohydramnion, te weinig vruchtwater.
- Pre-eclampsie of zwangerschapsvergiftiging
- Stuitligging
- Vroeggeboorte
- Vanishing Twin Syndrome, het verdwijnen van een van de twee embryo's
- Vroegtijdige en voortijdige gebroken vliezen
- Vroegtijdige en voortijdige weeën
- Zwangerschapsmisselijkheid of hyperemesis gravidarum
Behalve bovenstaande problemen en ziekten, zijn er ook problemen die tijdens zwangerschap ontstaan of verergeren, maar die ook voorkomen zonder zwangerschap. Dat zijn bijvoorbeeld suikerziekte, schildklierziekten: hyperthyreoïdie en hypothyreoïdie, stollingsstoornissen.
Buitenbaarmoederlijke zwangerschap[bewerken | brontekst bewerken]
Normaal nestelt de bevruchte eicel zich in de baarmoeder of uterus. Nestelt de eicel zich ergens anders, dan spreekt men van een buitenbaarmoederlijke zwangerschap of 'extra-uteriene graviditeit'.
Begeleiding[bewerken | brontekst bewerken]
Roken en alcohol[bewerken | brontekst bewerken]
Zowel roken als het gebruik van alcohol tijdens de zwangerschap kunnen nadelige gevolgen hebben voor de gezondheid en ontwikkeling van het ongeboren kind. Toch blijft één op de vier vrouwen drinken en blijft ongeveer één op de vijf zwangere vrouwen roken.[4][5][bron?]
Alcohol[bewerken | brontekst bewerken]
Alcohol dringt door de placenta en kan daardoor de foetus beschadigen in alle stadia van de zwangerschap. Kinderen die zijn blootgesteld aan alcohol tijdens de zwangerschap huilen meer, slapen slechter, reageren trager en kalmeren ook langzamer na opwinding. Ze hebben gemiddeld een lager IQ, meer leerstoornissen en sociale problemen. Zelfs consumptie van minder dan één glas alcohol per dag kan leiden tot een verhoogde kans op groeiafwijkingen, een miskraam en doodgeboorte.[6]
Blootstelling aan dagelijkse hoge alcoholdoseringen tijdens de ontwikkeling van de foetus kan leiden tot het foetale-alcoholsyndroom (FAS). FAS-kinderen hebben een groeiachterstand, afwijkende gelaatskenmerken en neurologische afwijkingen. Gerelateerde aandoeningen zijn Alcohol-Related Neurodevelopmental Disorder (ARND) en Alcohol-Related Birth Defects (ARBD).[6] ARND lijkt op FAS, maar dan zonder de uiterlijke kenmerken en ARBD betreft lichamelijke afwijkingen zoals hart-, bot- en orgaanproblemen.
Roken[bewerken | brontekst bewerken]
Giftige stoffen die worden geïnhaleerd door roken of meeroken, zoals nicotine, cadmium en koolmonoxide, beïnvloeden de groei van de vrucht. Nicotine leidt tot vaatvernauwing en koolmonoxide neemt in het bloed de plaats in van zuurstof.[5] Daardoor krijgt de vrucht minder bloed, dus minder voedingsstoffen, en minder zuurstof. Mede daardoor is het geboortegewicht van de baby van rokende vrouwen gemiddeld 250 gram lager. Een lager geboortegewicht geeft ook een grotere kans op perinatale sterfte van het kind. Roken leidt ook tot verhoogde kans op onvruchtbaarheid: bij rokende vrouwen is de tijd tot de conceptie gemiddeld 30% langer dan bij niet-rokende vrouwen. Roken tijdens de zwangerschap vergroot de kans op een miskraam of vroeggeboorte. In het eerste levensjaar van de baby is de kans op luchtweginfecties en wiegendood groter.
Cadmium veroorzaakt dikkere basaalmembranen om kleine bloedvaten, in de gonaden en ook in de placenta, bijgevolg wordt de uitwisseling van voedingstoffen en het metabolisme voor de foetus bemoeilijkt. Door de schade in de gonaden zijn er minder gezonde sperma- en ei-cellen.
Zelfs de kleinkinderen van vrouwen die roken tijdens de zwangerschap kunnen hier nog schade van ondervinden, namelijk een hogere kans op astma en een slechtere longfunctie. Dit geldt vrijwel uitsluitend voor de kleinzoons, van wie de oma aan de kant van hun moeder rookte tijdens de zwangerschap. De relatie van grootmoeder via de moeder hangt samen met het feit dat eicellen in een vrouwelijk embryo al tijdens de zwangerschap worden aangelegd. Zaadcellen ontstaan pas in en na de puberteit.[7]
Voeding[bewerken | brontekst bewerken]
Het Voedingscentrum heeft een lijst van voedingsmiddelen opgesteld die vrouwen tijdens de zwangerschap beter niet kunnen eten met onder andere rauw vlees, rauwmelkse kaas, roofvissen, geneeskrachtige kruiden, drop.[8] Tijdens de zwangerschap kunnen de eetgewoonte drastisch veranderen. De drang naar zoet, en soms weer zuur voedsel wordt groter. Uit onderzoek van de Queens Mary Universiteit is gebleken dat vrouwen die tijdens hun zwangerschap veel suikerhoudende producten eten, meer kans hebben dat het kind te kampen krijgt luchtwegproblemen, zoals astma en stofallergie.[9] Een ander gevolg van deze verandering is dat deze toename van zoete en/of zure producten een negatieve impact kan hebben op het gebit. Dit heeft als gevolg dat het tandglazuur aangetast kan worden. Dit verhoogt de risico op gaatjes en tanderosie. Experts in tandheelkunde geven dan ook aan dat het van uiterst belang is dat er extra aandacht wordt besteed aan de verzorging van het gebit tijdens de zwangerschap.[10]
Vrijen[bewerken | brontekst bewerken]
Tijdens het eerste trimester van de zwangerschap kan de vrouw last hebben van misselijkheid, braakneigingen of vermoeidheid. Deze factoren kunnen ervoor zorgen dat de zin of het plezier in vrijen vermindert. Ook moeten sommige vrouwen vaker plassen, een gevoel dat tijdens geslachtsgemeenschap nog kan worden versterkt. Het feit dat de borsten gevoeliger worden, maakt dat sommige vrouwen het aanraken ervan onprettig vinden. Anderen vinden juist seksueel genot in deze verhoogde gevoeligheid.
Veel van de ongemakken uit het begin van de zwangerschap, verdwijnen in het tweede trimester. De vagina wordt breder en produceert meer vocht, dat lichtjes van geur verschilt ten opzichte van vroeger. Veel vrouwen voelen zich in deze periode sneller of vaker opgewonden.[bron?]
Het is een wijdverbreid misverstand dat vrijen tijdens de laatste fase, het derde trimester van de zwangerschap aanleiding zou kunnen geven tot een te vroege geboorte. Uit onlangs verschenen rapporten blijkt dat deze veronderstelling onjuist is.[bron?] In gevallen dat vrijen een oorzaak leek te zijn van een vervroegde bevalling, bleek de werkelijke oorzaak vaak een infectie in de vagina te zijn. Als er geen sprake was van ziekmakende micro-organismen in de vagina, leek eerder het tegendeel waar te zijn: meer vrijen leidde dan juist tot minder vroeggeboorten.
Het vrijen in de missionarishouding, waarbij de man op de vrouw ligt, kan echter wel door de dikke buik bemoeilijkt worden. Borststimulatie kan resulteren in de afscheiding van biestmelk of colostrum, wat volkomen normaal is maar door sommige koppels als storend wordt ervaren. Soms raadt de arts geslachtsverkeer af. Dit gebeurt meestal indien er zich ernstige problemen voordoen tijdens de zwangerschap.
Zie ook[bewerken | brontekst bewerken]
- Chronologie van de menselijke zwangerschapsontwikkeling
- Couvade
- Echografie bij zwangerschap
- Zwangerschapscursus
- Zwangerschapsdiscriminatie
Externe link[bewerken | brontekst bewerken]
Zwangerschap in kunst[bewerken | brontekst bewerken]
-
 Zwangerschapskorset
Zwangerschapskorset -
 Een zwangere vrouw,
Een zwangere vrouw,
18e eeuw -
 Beeld van een zwangere vrouw,
Beeld van een zwangere vrouw,
Danny Osborne -
 Man en zwangere vrouw,
Man en zwangere vrouw,
Gustav Klimt -
 Schilderij,
Schilderij,
Gustav Klimt, 1903 -
 Zwangere vrouw met kind,
Zwangere vrouw met kind,
Heinrich Zille -
 Twee handen,
Twee handen,
Guus Hellegers, 1986 -

-
 Cycladenidool van een zwangere vrouw
Cycladenidool van een zwangere vrouw -
 Cycladenidool van een zwangere vrouw
Cycladenidool van een zwangere vrouw -
 The Pregnant Woman,
The Pregnant Woman,
Toronto -
 Beeld van zwangere vrouw
Beeld van zwangere vrouw -
 Tempel van Hatsjepsoet
Tempel van Hatsjepsoet -

-
 Zwangere vrouw gekleed in de zon staat op de maan, 1450
Zwangere vrouw gekleed in de zon staat op de maan, 1450 -
 Zwangere vrouw en man
Zwangere vrouw en man -
 Beeld van zwangere vrouw, Mexico
Beeld van zwangere vrouw, Mexico -
 Zwangere vrouw van Steven Whyte
Zwangere vrouw van Steven Whyte












.jpg)


