
Gang (dieren)
Gang is het bewegingspatroon van de ledematen van dieren tijdens voortbeweging over een oppervlak. De meeste dieren gebruiken verschillende gangen, afhankelijk van snelheid, terrein, noodzaak tot manoeuvreren, stabiliteit en efficiënt energieverbruik. Verschillende dieren hebben verschillende gangen door verschillen in anatomie of aangeboren voorkeur. Pas sinds de chronofotografie van Eadweard Muybridge en Étienne-Jules Marey kan er goed onderzoek naar de verschillende gangen worden gedaan. Milton Hildebrand was een pionier op het gebied van de wetenschappelijke analyse en classificatie van de gangen, terwijl Luis de Ascásubi in dezelfde periode tot een classificatie kwam.
Er zijn vele verschillende soorten gangen, waarbij drie algemene klasses te onderscheiden zijn: stap, draf en galop. Bij de paardengang zijn tot wel 60 verschillende patronen gevonden, met 37 stap, 14 draf en 9 galop.[1] Met het blote oog zijn deze moeilijk te onderscheiden.
Er zijn verschillende criteria om de diverse gangen te onderscheiden, waaronder symmetrie, de faseverhouding tussen voor- en achterledemaat en de verhouding tussen stand- en zwaaifase, de duty-factor.

Een gang is symmetrisch als alle ledematen in dezelfde mate worden gebruikt. Zo zijn lopen en draven symmetrisch en zijn gallop en sprong asymmetrisch.
Daarnaast kan onderscheid gemaakt worden tussen laterale en diagonale paren. Bij een lateraal paar worden de benen aan dezelfde zijde vrijwel gelijktijdig verzet. Bij diagonaal paar is dit kruislings.
De volgorde waarin benen verzet worden, kan ook lateraal of diagonaal zijn, waarbij de benaming afhankelijk is van welk been als referentie wordt gekozen. Zo is stap lateraal als het linkerachterbeen de referentie is, maar diagonaal als het linkervoorbeen dat is.
Bij rennende gangen maakt de voet minder dan de helft van de tijd contact met de grond.
Symmetrisch[bewerken | brontekst bewerken]

Bekende gangen zijn benoemd: rentelgang (pace), paso, stap (walk), langzame tölt (slow gait), snelle tölt (rack), snelstap (running walk), langzame draf en draf. Daartussen vallen de ongewone gangen

Hildebrand publiceerde in 1965 een studie van symmetrische gangen waarin hij film had geanalyseerd van 64 en 128 beelden per seconde. Hij kwam zo tot gangformules met daarin de volgorde van de passen, de paslengte en -tijd en de stand- en zwaaitijd. Dit waren middelingen van waarnemingen van verschillende paarden, aangezien er altijd enige variatie is. Symmetrische gangen kunnen worden uitgedrukt met twee variabelen als de contactinterval voor en achter gelijk is:
- de standfase van het achterbeen
- de pasinterval tussen stap van het achter- en het voorbeen aan dezelfde zijde
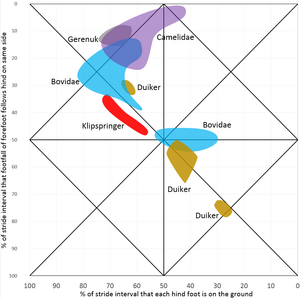
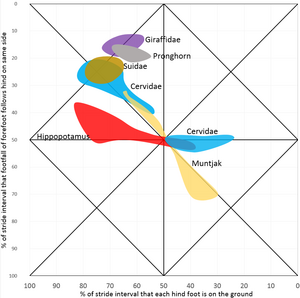
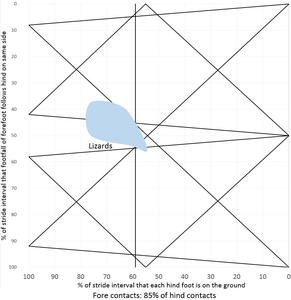
De eerste variabele geeft aan hoeveel tijd de voeten op de grond staan en de tweede variabele vergelijkt de voorvoeten als paar met de achtervoeten als paar. Dit kan worden uitgezet in een grafiek zoals hiernaast met de eerste variabele op de x-as en de tweede op de y-as. De x-as kent twee absolute grenzen, aangezien de voeten aan de linkerzijde nooit de grond verlaten en aan de rechterzijde nooit de grond raken. De boven- en onderzijde zijn gelijk aan elkaar, een verschil in pasinterval van achter- en voorbeen aan dezelfde zijde van 0% is gelijk aan 100% en tussen 3% en 97% zit dus maar 6% verschil. De grafiek zou opgerold kunnen worden als cilinder die raakt aan boven- en onderzijde.
In het diagram liggen vergelijkbare patronen dichtbij elkaar en kan de bijbehorende gang benoemd worden. Daarbij sloot Hildebrand zoveel mogelijk aan bij bestaande namen, maar dit was niet altijd mogelijk, omdat niet alle mogelijke gangen benoemd zijn en sommige namen voor verschillende gangen gebruikt worden.
Als de contactinterval voor en achter niet gelijk is, dan kan de gang uitgedrukt worden met een derde variabele:
- de verhouding van de standfase van het voorbeen ten opzichte van het achterbeen
Deze derde variabele zou in een driedimensionale grafiek uitgezet kunnen worden, maar Hildebrand koos voor een overlay.
Ten tijde van de studie uit 1965 waren er 1008 mogelijke gangformules berekend voor 158 geslachten van viervoetigen, waaronder 8 voor amfibieën, 9 voor reptielen en 15 ordes van zoogdieren. Voor het paard zijn 167 formules van toepassing. In 1976 gebruikte Hildebrand 1178 formules.
Geen van de geanalyseerde dieren plaatst de voorvoet vlak voor het plaatsen van de achtervoet.
Rentelgang is een fractie sneller dan draf en heeft als voordeel dat voor- en achtervoet elkaar niet kunnen raken.
Voor de symmetrische gangen zijn er 16 basisformules en 28 overgangsformules, in totaal 44. Zoals in de grafiek is te zien, zijn er binnen een gang variaties. Zo gaf Hildebrand de volgende mogelijkheden weer, waarbij ⚫ de standfase is en ⚪ de zwaaifase, L left, R right, H hind foot en F fore foot. Een deel van deze gangen is eigenlijk asymmetrisch.
| Rentelgang | ||||
|---|---|---|---|---|
| 10 | 57 | 35 | ||
| LH | ⚫⚫⚪⚪⚪⚪⚪⚪ | ⚫⚫⚫⚪⚪⚪⚪⚪ | ⚫⚪⚪⚪ | ⚫⚫⚪⚪⚪⚪ |
| LF | ⚪⚫⚫⚪⚪⚪⚪⚪ | ⚪⚫⚪⚪⚪⚪⚪⚪ | ⚫⚪⚪⚪ | ⚪⚫⚪⚪⚪⚪ |
| RF | ⚪⚪⚪⚪⚪⚫⚫⚪ | ⚪⚪⚪⚪⚪⚫⚪⚪ | ⚪⚪⚫⚪ | ⚪⚪⚪⚪⚫⚪ |
| RH | ⚪⚪⚪⚪⚫⚫⚪⚪ | ⚪⚪⚪⚪⚫⚫⚫⚪ | ⚪⚪⚫⚪ | ⚪⚪⚪⚫⚫⚪ |
| Stap | ||||
| 4 | 22 | 3 | 20 | |
| LH | ⚫⚫⚫⚫⚫⚪⚪⚪ | ⚫⚫⚫⚫⚪⚪ | ⚫⚫⚫⚫⚫⚫⚪⚪ | ⚫⚫⚫⚪ |
| LF | ⚪⚪⚫⚫⚫⚫⚫⚪ | ⚪⚪⚫⚫⚫⚫ | ⚫⚪⚪⚫⚫⚫⚫⚫ | ⚪⚫⚫⚫ |
| RF | ⚫⚫⚫⚪⚪⚪⚫⚫ | ⚫⚫⚫⚪⚪⚫ | ⚫⚫⚫⚫⚫⚪⚪⚫ | ⚫⚫⚪⚫ |
| RH | ⚫⚪⚪⚪⚫⚫⚫⚫ | ⚫⚪⚪⚫⚫⚫ | ⚫⚫⚪⚪⚫⚫⚫⚫ | ⚫⚪⚫⚫ |
| Paso, tölt, snelstap | ||||
| 9 | 50 | |||
| LH | ⚫⚫⚫⚪⚪⚪⚪⚪ | ⚫⚫⚫⚫⚫⚪⚪⚪ | ⚫⚫⚫⚪⚪⚪ | ⚫⚫⚫⚫⚪⚪ |
| LF | ⚪⚪⚫⚫⚫⚪⚪⚪ | ⚪⚪⚪⚫⚫⚫⚪⚪ | ⚪⚪⚫⚫⚪⚪ | ⚪⚪⚫⚫⚫⚪ |
| RF | ⚫⚪⚪⚪⚪⚪⚫⚫ | ⚫⚫⚪⚪⚪⚪⚪⚫ | ⚫⚪⚪⚪⚪⚫ | ⚫⚫⚪⚪⚪⚫ |
| RH | ⚪⚪⚪⚪⚫⚫⚫⚪ | ⚫⚪⚪⚪⚫⚫⚫⚫ | ⚪⚪⚪⚫⚫⚫ | ⚫⚪⚪⚫⚫⚫ |
| Draf | ||||
| 11 | 14 | 58 | 41 | |
| LH | ⚫⚫⚪⚪⚪⚪⚪⚪ | ⚫⚫⚪⚪⚪⚪⚪⚪ | ⚫⚫⚫⚪⚪⚪⚪⚪ | ⚫⚪⚪⚪ |
| LF | ⚪⚪⚪⚫⚫⚪⚪⚪ | ⚪⚪⚪⚪⚪⚫⚫⚪ | ⚪⚪⚪⚪⚪⚫⚪⚪ | ⚪⚪⚫⚪ |
| RF | ⚫⚪⚪⚪⚪⚪⚪⚫ | ⚪⚫⚫⚪⚪⚪⚪⚪ | ⚪⚫⚪⚪⚪⚪⚪⚪ | ⚫⚪⚪⚪ |
| RH | ⚪⚪⚪⚪⚫⚫⚪⚪ | ⚪⚪⚪⚪⚫⚫⚪⚪ | ⚪⚪⚪⚪⚫⚫⚫⚪ | ⚪⚪⚫⚪ |
Verschillen tussen de soorten[bewerken | brontekst bewerken]
Elke diersoort gebruikt slechts een beperkt aantal van de beschikbare gangen. Vrijwel alle lopende dieren beheersen symmetrische gangen, terwijl asymmetrische gangen vooral door zoogdieren gebruikt worden.
.jpg)
De meeste viervoetigen gebruiken laterale gangen, draf of rentelgang, maar arboreale dieren als de meeste primaten, opossums en kinkajoes lopen met een diagonale gang. De huisbuidelspitsmuis, een opossum, leeft overwegend op de grond en heeft dan ook vooral een laterale gang, terwijl een vooral in bomende levende opossum, de gele wolhaarbuidelrat, vooral een diagonale gang heeft. Hierbij lijkt het een rol te spelen dat arboreale dieren veel op smalle ondersteuningen lopen.[2]
-
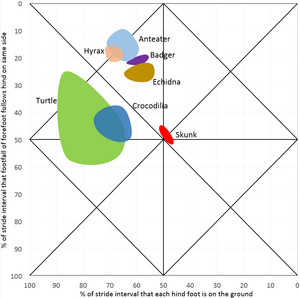
-
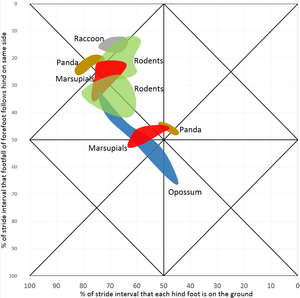 wasbeer (Procyon lotor), panda Ailurus, knaagdieren boven Cavia, Dolichotis, Rattus, Cynomys, Hydrochoerus, knaagdieren onder Marmota, Castor, Dipodomys, Dasyprocta, buideldieren boven Dendrolagus, Sarcophilus, Phascolomis, Petaurus, buideldieren onder Marmosa, Lasiorhinus, Petaurus, opossum Didelphis
wasbeer (Procyon lotor), panda Ailurus, knaagdieren boven Cavia, Dolichotis, Rattus, Cynomys, Hydrochoerus, knaagdieren onder Marmota, Castor, Dipodomys, Dasyprocta, buideldieren boven Dendrolagus, Sarcophilus, Phascolomis, Petaurus, buideldieren onder Marmosa, Lasiorhinus, Petaurus, opossum Didelphis -
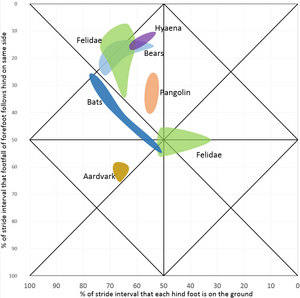 hyena Hyaena, katachtigen Felis, Panthera, Acinonyx, beren Thalarctos, Melursus, Helarctos, Tremarctos, Selenarctos, Ursus, schubdierachtige Manis, vleermuizen Pteropus, Tadarida, aardvarken Orycteropus
hyena Hyaena, katachtigen Felis, Panthera, Acinonyx, beren Thalarctos, Melursus, Helarctos, Tremarctos, Selenarctos, Ursus, schubdierachtige Manis, vleermuizen Pteropus, Tadarida, aardvarken Orycteropus -
 civetkatachtigen Arctictis, Paguma, Crossarchus, Suricata, Atilax, Ichneumia, Nandinia, Herpestes, wasbeerhond Nyctereutes, hondachtigen Lycaon, Urocyon, gordeldier boven Dasypus, gordeldier beneden Priodontes, egel Hemiechinus, tenrek Echinops, kinkajoe Potos
civetkatachtigen Arctictis, Paguma, Crossarchus, Suricata, Atilax, Ichneumia, Nandinia, Herpestes, wasbeerhond Nyctereutes, hondachtigen Lycaon, Urocyon, gordeldier boven Dasypus, gordeldier beneden Priodontes, egel Hemiechinus, tenrek Echinops, kinkajoe Potos -
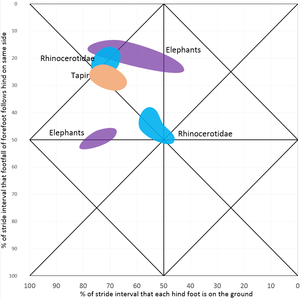
-
-
-

-

-
-












Ademhaling[bewerken | brontekst bewerken]

Bij de vroegste viervoeters werd mogelijk de ademhaling belemmerd door een gang die gekenmerkt wordt door een spreidstand van het heupgewricht en een zijdelingse wervelbuiging. De huidige hagedissen hebben een vergelijkbare anatomie waarbij dit het geval is, doordat spieren van de ribbenkast zowel voor de voortbeweging als voor de ademhaling worden gebruikt. Veel koudbloedigen hebben deze beperking – door Richard Cowen naar David Carrier Carrier's constraint genoemd – en vertrouwen daarom op anaerobe verbranding voor korte uitbarstingen, of hebben passieve verdedigingsmechanismes. In de evolutie van warmbloedige zoogdieren is dit gebrek aan uithoudingsvermogen ondervangen, terwijl ook vogels lange tijd grote inspanningen kunnen leveren. Ook vissen als de ruwe haai en de witte tonijn kunnen zich dagenlang voortbewegen. Amfibieën weten de beperking te omzeilen met keelademhaling en huidademhaling. Enkele hagedissen als de varaan maken ook gebruik van keelademhaling.
De zijdelingse beweging bij hagedissen vereist dat de spieren aan de ene zijde samentrekken en aan de andere zijde ontspannen, terwijl bij de ademhaling de beide zijden gelijktijdig samentrekken en ontspannen. Daarnaast helpt laterale buiging niet om lucht in en uit beide longen te pompen, doordat de buigingsas door het midden van de romp loopt. Waar aan de ene zijde het volume vergroot wordt, wordt dat aan de andere zijde juist verkleind. Verder maakt de spreidstand van het heupgewricht dat er een laterale kracht op de romp komt te staan, waartegen spieren zich aanspannen die ook voor de ademhaling gebruikt worden.
.jpg)

In contrast met de laterale buiging bij hagedissen helpt de sagittale buiging bij zoogdieren in draaiende galop juist bij de ademhaling. Met de benen ver uit elkaar en een holle rug wordt de lucht aangezogen, terwijl de lucht uit de longen wordt gepompt bij de kromme rug als de benen onder het lichaam komen.[3] Een nadeel hiervan is dat de ademhaling niet sneller kan dan het looptempo, wat een limiet stelt aan de zuurstofhoeveelheid en de warmte-afvoer. Gangen waarbij geen sagittale buiging optreedt, zoals bij een hardlopende mens, zijn niet gelimiteerd door het looptempo.
Voorkeursgang[bewerken | brontekst bewerken]
Energie[bewerken | brontekst bewerken]
Elke gang kent een snelheid waarbij het energieverbruik per afgelegde afstand minimaal is. Om een minimaal energieverbruik te bereiken:
- zijn paslengte en -frequentie op elkaar afgestemd
- is de longventilatie afgestemd op de metabole zuurstofbehoefte
- zijn ademfrequentie en -volume op elkaar afgestemd; voor elke zuurstofbehoefte is er een optimum door de elastische weerstand van de borstkas, de weerstand van de luchtwegen en de dode ruimte in de longen
- is de ademhaling synchroon met de loopbewegingen
Deze optimale snelheid blijkt ook de voorkeurssnelheid te zijn van paarden. Bij een hogere snelheid past een andere voorkeursgang.
Onderzoek waarbij het zuurstofverbruik van paarden op een loopband werd gemeten, liet zien dat bij een snelheid van 4,5 kilometer per uur het zuurstofverbruik van een dravend paard anderhalfmaal hoger lag dan wanneer het liep. Pas bij hogere snelheid werd het gunstiger om te draven.
Het minimum aan zuurstofverbruik lag vrijwel op hetzelfde niveau voor elke voorkeurssnelheid, zodat het voor het energieverbruik per afstand weinig uitmaakt of een paard een bepaalde afstand snel of langzaam aflegt, zolang de voorkeurssnelheid maar wordt aangehouden.[4] Ook bij migrerende dieren als de blauwe gnoe, de thomsongazelle en andere dieren werden voorkeurssnelheden vastgesteld.[5]
Optimale loopsnelheden zijn deels afhankelijk van de samentrekkende eigenschappen van de skeletspieren. Er is een optimale snelheid van spierverkorting en daarmee een optimale pasfrequentie. Daarnaast neemt het vermogen van spieren om kracht te genereren af naarmate de spier verder dan de optimale lengte wordt uitgerekt of ingekort en daarmee is er ook een optimale paslengte. Aangezien staplengte en stapfrequentie de snelheid bepalen, bepalen deze twee de optimale snelheid in alle gangen. De mens lijkt echter een uitzondering.
Bij de mens is er wel een verschil tussen het energieverbruik per afstand bij lopen en rennen. Bij de optimale snelheid verbruikt een lopend mens aanmerkelijk minder energie per afstand dan een rennend mens. Rennen kent bij de mens geen optimale snelheid, het energieverbruik per afstand neemt steeds verder af bij toenemende snelheid.[6]
Getal van Froude[bewerken | brontekst bewerken]
Robert McNeill Alexander kwam met een methode om de gang van dieren van verschillende grootte met elkaar te vergelijken. Hiertoe gebruikte hij het getal van Froude Fr:

waarbij:
- v: snelheid (m/s)
- g: gravitatie (m/s2)
- l: lengte van het achterbeen tot aan de heup
Daarbij wordt het been benaderd als omgekeerde slinger. Het froudegetal is kleiner dan 1 bij lopen voor tweevoetigen of stap voor viervoetigen, aangezien daarboven een zweefmoment ontstaat, maar de overgang naar rennen voor tweevoetigen en draf voor viervoetigen wordt veelal al gemaakt tussen 0,3 en 0,8. Om dichter bij 1 te komen, zou de paslengte af en de pasfrequentie toe moeten nemen tot voor lopen oncomfortabele hoogte. Tussen 2 en 3 wordt veelal de overgang naar galop gemaakt.[7]
Zespotigen[bewerken | brontekst bewerken]
Bij zespotigen zijn vele verschillende gangen mogelijk, maar de voorkeur heeft die waarbij de buitenste poten aan de ene kant en de middelste aan de andere worden verzet en daarna andersom. Daarbij staan steeds drie poten aan de grond, wat de stabiliteit ten goede komt, aangezien een driepoot statisch stabiel is.
Hoewel deze afwisselende driepootsgang veel voorkomt, kennen insecten net als gewervelden verschillende gangen voor verschillende snelheden. Zo gaat de bekende bananenvlieg over van een langzame golfbeweging naar een vierpootsgang en uiteindelijk de snelle tripod, al zijn de overgangen hiertussen minder abrupt dan bij gewervelden. Daarbij neemt de paslengte lineair toe. De vlieg houdt bij deze gangen steeds respectievelijk vijf, vier en drie poten aan de grond. Dit is op vlak terrein minder efficiënt dan de snellere gangen van gewervelden, die daarbij slechts een of twee voeten aan de grond hebben. De hypothese is lang geweest dat hoewel de driepootsgang langzamer is op vlak terrein, deze gang bij klimmen wel sneller is. Dit blijkt inderdaad het geval met poten die over adhesie beschikken. Zodra deze adhesie uitgeschakeld wordt, blijken de bananenvliegen over te gaan op een snellere tweevoetige gang die te vergelijken valt met draf.[8]
Wandelende takken en wandelende bladeren gebruiken een vierpootsgang bij langzaam lopen en gaan over naar een driepootsgang als ze versnellen. Kakkerlakken gebruiken twee gangen, een om langzaam en een om snel te lopen, maar beide zijn driepootsgangen.[9] Bij de kakkerlak Nauphoeta cinerea is echter waargenomen dat deze voor de snelle gang overgaat naar een metachrone golfbeweging, waarbij de passen van de ipsilaterale poten – de poten aan dezelfde zijde – elkaar opvolgen. Hierbij verminderen de verticale bewegingen, wat gunstig kan zijn voor dieren die maar beperkte mogelijkheden hebben om gebruik te maken van de elastische energie in de poten, zoals wel het geval is bij springende gangen als de draf. Ook lijkt deze gang geschikte voor gladde ondergronden.[10]
Ook tijdens het draaien wordt de driepootgang veelal aangehouden, wat algemeen voorkomt bij geleedpotigen. Aanpassing van de pasfrequentie ligt ook niet voor de hand, vooral niet bij brede dieren als kakkerlakken die de pasfrequentie aan een zijde bijna zouden moeten verdubbelen.[11]
De krachten per potenpaar verschillen veelal. Het borststuk is te verdelen in voor-, midden- en achterborst of pro-, meso- en metathorax. Bij krekels en sprinkhanen levert het prothoracisch potenpaar minder kracht en lijkt dit vooral gebruikt te worden om te balanceren.[12]
Literatuur[bewerken | brontekst bewerken]
- Hildebrand, M. (1965): 'Symmetrical Gaits of Horses' in Science, Volume 150, Issue 3697, p. 701-708
- Hildebrand, M. (1976): 'Analysis of tetrapod gaits: general considerations' in Herman, R.M.; Grillner, S.; Stein, P.S.G.; Stuart, D.G. Neural Control of Locomotion, Springer
Noten[bewerken | brontekst bewerken]
- ↑ Roberts, T.D.M. (1995): Understanding Balance. The Mechanics of Posture and Locomotion, Nelson Thornes, p. 211
- ↑ Lemelin, P.; Schmitt, D.; Cartmill, M. (2003): 'Footfall patterns and interlimb co-ordination in opossums (Family Didelphidae): evidence for the evolution of diagonal-sequence walking gaits in primates' in Journal of Zoology, Volume 260, Issue 4, p. 423-429
- ↑ Carrier, D.R. (1987): The evolution of locomotor stamina in tetrapods: circumventing a mechanical constraint' in Paleobiology, Volume 13, Issue 3, p. 326-341
- ↑ Hoyt, D.F.; Taylor, C.R. (1981): 'Gait and the energetic of locomotion in horses' in Nature, Volume 292, Issue 5820, p. 239-240
- ↑ Pennycuick, C.J. (1975): 'On the running of the gnu, Connochaetes taurinus, and other animals' in Journal of Experimental Biology, Volume 63, p. 775-99
- ↑ Carrier, D.R. (1984): 'The Energetic Paradox of Human Running and Hominid Evolution' in Current Anthropology, Volume 25, No. 4, p. 483-495
- ↑ Alexander, R.M. (2003): Principles of Animal Locomotion, Princeton University Press, p. 59
- ↑ Ramdya, P.; Thandiackal, R.; Cherney, R.; Asselborn, T.; Benton, R.; IJspeert, A.J.; Floreano, D. (2017): 'Climbing favours the tripod gait over alternative faster insect gaits' in Nature Communications, Volume 8
- ↑ Mendes, C.S.; Bartos, I.; Akay, T.; Márka, S.; Mann, R.S. (2013): 'Quantification of gait parameters in freely walking wild type and sensory deprived Drosophila melanogaster' in eLife. Gearchiveerd op 26 mei 2022.
- ↑ Weihmann, T.; Brun, P.G.; Pycroft, E. (2017): 'Speed dependent phase shifts and gait changes in cockroaches running on substrates of different slipperiness' in Frontiers in Zoology. Gearchiveerd op 12 juli 2023.
- ↑ Jindrich, D.; Full, R.J. (1999): 'Many-legged maneuverability: Dynamics of turning in hexapods' in Journal of Experimental Biology, Volume 202, p. 1603-1623
- ↑ Harris, J.; Ghiradella, H. (1980): 'The forces exerted on the substrate by walking and stationary crickets' in Journal of Experimental Biology, Volume 85, p. 263-279