
Rondwormen: verschil tussen versies
| Regel 179: | Regel 179: | ||
Andere rondwormen tasten bomen aan. De belangrijkste vertegenwoordiger van deze groep is ''[[Bursaphelenchus xylophilus]]'', de rondworm dat in [[Den (geslacht)|dennen]]bossen voorkomt in Azië en Amerika en recent ontdekt is in Europa. |
Andere rondwormen tasten bomen aan. De belangrijkste vertegenwoordiger van deze groep is ''[[Bursaphelenchus xylophilus]]'', de rondworm dat in [[Den (geslacht)|dennen]]bossen voorkomt in Azië en Amerika en recent ontdekt is in Europa. |
||
Parasitaire plantenaaltjes kunnen '''bestreden''' worden door [[vruchtwisseling]] of gebruik te maken van [[Resistentie bij planten|resistente]] rassen. Bij het gebruik van resistente rassen kunnen echter [[pathotype]]n gaan optreden, zoals bij het [[aardappelcystenaaltje]], waarvan thans 5 pathotypen bestaan. Het blijft dan een race tussen de [[Plantenveredeling|plantenverdelaar]] en de aanpassingssnelheid van het aaltje. Ook kunnen plantensoorten als ''[[Tagetes patula]]'' of andere ''[[Tagetes]]''-soorten ingezet worden tegen de [[wortellesieaaltjes]].<ref name="Riotte" /> Het wortelexudaat van de Tagetesplanten [[Thiofeen|thiophene a-terthienyl]] doodt de aaltjes.<ref>[https://www.gardentara.com/companion-planting-with-marigolds/ Companion planting with marigolds: myths and secrets, Published by Amber Hall on July 3, 2018]</ref> Een nadeel hiervan is dat er in het betreffende seizoen geen gewas geteeld kan worden. Een andere mogelijkheid is het gebruik van natuurlijke [[antagonist]]en zoals de schimmel ''[[Gliocladium roseum]]''. [[Chitosan]], een natuurlijk [[biocide]], lokt plantenafweerreacties uit waarbij parasitaire [[cystenaaltje]]s op de wortels van [[soja]], [[maïs]], [[suikerbiet]], [[aardappel]] en [[tomaat]] gedood worden, maar niet de niet-parasitaire aaltjes in de bodem.<ref name="US-2008/072494">{{ cite patent | country = US | number = 2008072494 | status = application | title = Micronutrient elicitor for treating nematodes in field crops | pubdate = 2008-03-27 | fdate = 2006-09-07 | inventor = Stoner RJ, Linden JC | assign1 = }}</ref> Stomen van de grond, waarbij de grondtemepratuur renminste moet oplopen tot 80°C, is een effectieve methode om aaltjes te doden voordat een gewas wordt geplant, maar doodt zowel de schadelijke als de nuttige [[bodemfauna]]. [[CSIRO]] heeft een 13- tot 14-voudige afname van de populatiedichtheden van ''[[Meloidogyne javanica]]''-aaltjes gevonden in percelen met het [[sareptamosterd]]ras Nemfix (''Brassica juncea'' var. ''juncea'') als [[Groenbemesting|groenbemester]], dat dient als [[vanggewas]] voor de aaltjes.<ref name="CSIRO" /> Door de wortels van het vanggewas worden de larven uit de cysten gelokt, maar ze kunnen zich hierop niet vermeerderen. Ook kan [[bladrammenas]] als groenbemester ingezet worden tegen het [[bietencystenaaltje]] en [[aardappelmoeheid]], maar bladrammenas is een [[waardplant]] van het [[wit bietencystenaaltje|witte]] (''Heterodera schachtii'') en het [[geel bietencystenaaltje|gele bietencystenaaltje]] (''Heterodera trifolii''). Daarom moeten er tegen deze aaltjes resistente rassen van bladrammenas worden gebruikt. |
Parasitaire plantenaaltjes kunnen '''bestreden''' worden door [[vruchtwisseling]] of gebruik te maken van [[Resistentie bij planten|resistente]] rassen. Bij het gebruik van resistente rassen kunnen echter [[pathotype]]n gaan optreden, zoals bij het [[aardappelcystenaaltje]], waarvan thans 5 pathotypen bestaan. Het blijft dan een race tussen de [[Plantenveredeling|plantenverdelaar]] en de aanpassingssnelheid van het aaltje. Ook kunnen plantensoorten als ''[[Tagetes patula]]'' of andere ''[[Tagetes]]''-soorten ingezet worden tegen de [[wortellesieaaltjes]].<ref name="Riotte" /> Het wortelexudaat van de Tagetesplanten [[Thiofeen|thiophene a-terthienyl]] doodt de aaltjes.<ref>[https://www.gardentara.com/companion-planting-with-marigolds/ Companion planting with marigolds: myths and secrets, Published by Amber Hall on July 3, 2018]</ref> Een nadeel hiervan is dat er in het betreffende seizoen geen gewas geteeld kan worden. Een andere mogelijkheid is het gebruik van natuurlijke [[antagonist]]en zoals de schimmel ''[[Gliocladium roseum]]'', waarvan de [[conidium|conidia]] zich hechten aan de cuticula van het aaltje, ontkiemen en met hun kiemhyfen het aaltje binnendringen en doden.<ref name="green glow">{{cite journal|author=Zhang|display-authors=et al|year=2008|title=Investigation on the infection mechanism of the fungus ''Clonostachys rosea'' against nematodes using the green fluorescent protein|journal=Applied Microbiology and Biotechnology|volume=78|issue=6|doi=10.1007/s00253-008-1392-7|pages=983–990|pmid=18292995}}</ref> [[Chitosan]], een natuurlijk [[biocide]], lokt plantenafweerreacties uit waarbij parasitaire [[cystenaaltje]]s op de wortels van [[soja]], [[maïs]], [[suikerbiet]], [[aardappel]] en [[tomaat]] gedood worden, maar niet de niet-parasitaire aaltjes in de bodem.<ref name="US-2008/072494">{{ cite patent | country = US | number = 2008072494 | status = application | title = Micronutrient elicitor for treating nematodes in field crops | pubdate = 2008-03-27 | fdate = 2006-09-07 | inventor = Stoner RJ, Linden JC | assign1 = }}</ref> Stomen van de grond, waarbij de grondtemepratuur renminste moet oplopen tot 80°C, is een effectieve methode om aaltjes te doden voordat een gewas wordt geplant, maar doodt zowel de schadelijke als de nuttige [[bodemfauna]]. [[CSIRO]] heeft een 13- tot 14-voudige afname van de populatiedichtheden van ''[[Meloidogyne javanica]]''-aaltjes gevonden in percelen met het [[sareptamosterd]]ras Nemfix (''Brassica juncea'' var. ''juncea'') als [[Groenbemesting|groenbemester]], dat dient als [[vanggewas]] voor de aaltjes.<ref name="CSIRO" /> Door de wortels van het vanggewas worden de larven uit de cysten gelokt, maar ze kunnen zich hierop niet vermeerderen. Ook kan [[bladrammenas]] als groenbemester ingezet worden tegen het [[bietencystenaaltje]] en [[aardappelmoeheid]], maar bladrammenas is een [[waardplant]] van het [[wit bietencystenaaltje|witte]] (''Heterodera schachtii'') en het [[geel bietencystenaaltje|gele bietencystenaaltje]] (''Heterodera trifolii''). Daarom moeten er tegen deze aaltjes resistente rassen van bladrammenas worden gebruikt. |
||
Parasitaire plantenaaltjes kunnen ook bestreden worden door aaltjesetende aaltjes. |
Parasitaire plantenaaltjes kunnen ook bestreden worden door aaltjesetende aaltjes. |
||
Versie van 7 aug 2019 20:13
| Mee bezig Aan dit artikel of deze sectie wordt de komende uren of dagen nog druk gewerkt.
Klik op geschiedenis voor de laatste ontwikkelingen. |
| Rondwormen | |||||||
|---|---|---|---|---|---|---|---|

| |||||||
| Een rondworm die soja-bonen aantast (met ei). | |||||||
| Taxonomische indeling | |||||||
| |||||||
| Stam | |||||||
| Nematoda Rudolphi, 1808 | |||||||
| Afbeeldingen op | |||||||
| Rondwormen op | |||||||
| |||||||



Rondwormen (Nematoda) zijn een grote groep (stam) van zeer algemeen voorkomende wormen. Er zijn meer dan 25.000 beschreven soorten. Ook de aaltjes behoren tot de rondwormen. Plantenparasitaire rondwormen worden meestal aaltjes genoemd. Nematologie is de wetenschap die de nematoden bestudeert. De term nematoda komt uit het Grieks en betekent “draad-achtig”.
Veel rondwormen leven in de bodem van dode organische stof of eten bacteriën, schimmels en protozoa. Andere soorten leven als parasiet en veroorzaken plantenziekten zoals aardappelmoeheid, tasten insecten, naaktslakken aan of zitten in het darmkanaal van zoogdieren zoals de spoelwormen of de aarswormpjes en worden in het laatste geval dan vaak als draadworm aangeduid. Rondwormen worden ingezet ter bestrijding van schadelijke insecten,[1] zoals de inzet van Steinernema feltiae bij de bestrijding van de eikenprocessierups.[2] en het gebruik van Phasmarhabditis hermaphrodita tegen naaktslakken.
Sommige rondwormen kunnen wel 8 meter lang worden (Placentonema gigantissima),[3][4] maar de meeste rondwormen zijn voor het blote oog onzichtbaar en alleen door een microscoop zichtbaar. Ze zijn dan 5 tot 100 µm dik en 0,1 tot 2,5 mm lang.[5]. Per m² bodem kunnen wel een miljoen rondwormen leven. Ze komen overal voor.
Rondwormen kunnen aangetast worden door onder andere nematofage schimmels, virussen, sporozoën, amoeben, ciliaten, tardigraden, colembolen, oligochaeten en bacteriën.[6]
De soort Caenorhabditis elegans wordt gebruikt als modelorganisme. Het was de eerste meercellige eukaryoot waarvan het genoom volledig in kaart werd gebracht.[7] Ook is van C. elegans de ontwikkelingsbiologie van elke cel bepaald en elke neuron in kaart gebracht. Ook is het genoom van Meloidogyne incognita in 2008 volledig gesequenced.[8] Het genoom van Trichinella spiralis is eveneens volledig in kaart gebracht en in 2011 gepubliceerd.[9]
Lichaamsbouw
Rondwormen zijn ongesegmenteerde, ronde, oermondige dieren met een compleet spijsverteringsstelsel. Ze behoren daarmee tot de meest eenvoudige dieren die toch een compleet verteringsstelsel hebben. Ze hebben geen hart- en vaatstelsel of ademhalingsstelsel en zijn dus aangewezen op diffusie om zuurstof binnen te krijgen en te transporteren.[10]
De dwarsdoorsnede van rondwormen is rond en bestaat uit een kop, middenstuk en staart. De kop is radiaal symmetrisch en de rest van het lichaam is bilateraal symmetrisch.
De kop heeft in veel gevallen uit een geheel bestaande 'kopschermen' die rond de mond naar buiten uitstralen.[11] Bij de basis van de lippen zitten amfiden. Dit zijn de belangrijkste reukorganen van een rondworm. Elke amfide bestaat uit 12 sensorische zenuwcellen en met trilhaartjes bezette dendrieten.
Op het lichaam komen vaak groeven, ringen, borstelharen of andere onderscheidende structuren, zoals striae voor.[12]
Bij de geslachten Trichuris en Capillaria komen bacillaire banden voor. Dit zijn gespecialiseerde rijen van in de lengterichting liggende cellen. Ze bestaan uit kliercellen en een ander type cellen en worden gevormd door de hypodermis. De kliercellen hebben poriën tot in de huid. De functie van de bacillaire banden is onbekend. Hun buitengewone structuur duidt erop dat de kliercellen mogelijk een rol spelen in osmotische of ionenregulatie. De andere cellen spelen mogelijk een rol bij de vorming van de huid en de opslag van voedsel.
De 'huid' van een rondworm bestaat uit een syncytium of een enkele laag cellen en is bedekt met een dikke collagene cuticula of cuticulum. De cuticula heeft vaak een complexe structuur en kan uit twee of drie verschillende lagen bestaan. Onder de 'huid' ligt een laag in de lengte liggende spiercellen. De relatief stijve cuticula werkt samen met de spieren voor het vormen van een hydroskelet, omdat rondwormen geen spieren in de omtrek hebben. Zenuwuitsteeksels lopen vanuit het binnenste oppervlak van de spiercellen naar de aan de buikzijde gelegen zenuwbaan. Dit is uniek in het dierenrijk, waarbij de zenuwcellen uitlopers hebben naar de spieren in plaats van anders om.[11]
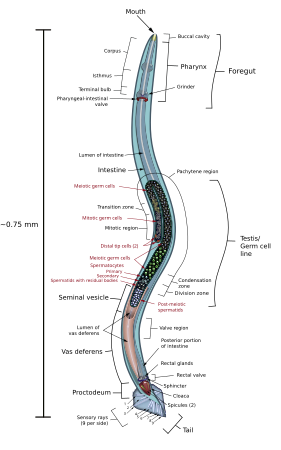
Een mannelijke rondworm bestaat van voor naar achteren uit een mondopening, darm (spijsverteringskanaal), cloaca, excretie-orgaan, testikel (mannelijk orgaan) en een excretie-opening. Aan de staartzijde kunnen 1 of 2 spicula, die dienen om de vulva van vrouwelijke rondwormen te openen en de overdracht van sperma te vergemakkelijken, hoewel sperma niet rechtstreeks door of met de spicula wordt overgedragen.[13][14] Het gubernaculum is een ander orgaan van het mannelijk copulatiesysteem, dat de spiculum of de spicula tijdens de copulatie geleidt. Alleen Dichelyne alatae heeft aan de staartzijde terhoogte van de cloaca-opening korte, brede vleugels, gevleugelde, ongelijke spicula en een schildvormig gubernaculum.
-
 13: Subventraal aanzicht van het voorste uiteinde van Torrestrongylus tetradorsalis met 'kopschermen', 14: Frontaalaanzicht met mondopening, papillen en amfiden.
13: Subventraal aanzicht van het voorste uiteinde van Torrestrongylus tetradorsalis met 'kopschermen', 14: Frontaalaanzicht met mondopening, papillen en amfiden. -
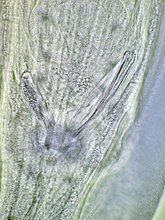
 Mannelijk staarteind van Capillaria plectropomi met de spinose spiculumschede en de uitgeschoven spiculum.
Mannelijk staarteind van Capillaria plectropomi met de spinose spiculumschede en de uitgeschoven spiculum. -
 Staarteind van de mannelijke rondworm Gongylonema pulchrum met rechts het spiculum en links het gubernaculum.
Staarteind van de mannelijke rondworm Gongylonema pulchrum met rechts het spiculum en links het gubernaculum. -
 Tekeningen van een mannelijke Philometra-rondworm - E en F zijn het gubernaculum.
Tekeningen van een mannelijke Philometra-rondworm - E en F zijn het gubernaculum.




Een vrouwelijke rondworm heeft als vrouwelijke organen een eierstok met een eileider, twee spermazakjes, baarmoeder en een vulva. Rondwormen hebben een simpel zenuwstelsel dat uit twee banen bestaat. De zenuwbaan aan de rugzijde (dorsaal) is groter dan die aan de buikzijde (ventraal). De strengen worden aan de voorkant om de farynx en soms ook richting het uiteinde met elkaar verbonden door dwarsverbindingen.[15] De lichaamsholte is een pseudocoeloom waarin vocht zit, dat helpt voedingsstoffen en zuurstof te verspreiden. Hun huid is bedekt met een cuticulum dat tegen uitdrogen beschermt. Hoewel dit cuticulum beweging en beperkte vormveranderingen toestaat groeit het niet mee en rondwormen moeten dan ook als ze groeien regelmatig vervellen. Er zijn vijf larvale stadia, L1, L2, L3 en L4. L5 is het volwassen stadium. In het ei vinden twee vervellingen plaats.
Mond
.jpg)
De mond heeft drie of zes lippen, die vaak een reeks tanden aan hun binnenranden dragen. In de mondholte kan een al of niet holle stilet voorkomen, waarmee de rondworm een cel kan aanprikken om deze vervolgens met de holle stilet leeg te zuigen. De stilet is een geharde, uitstekende opening van de darm (spijsverteringskanaal). Aan de basis van de stilet zitten twee knobbels.[16] Plantenparasitaire aaltjes, zoals Pratylenchus-soorten hebben een stilet.
Spijsverteringskanaal
De mondholte is bekleed met de cuticula, die vaak wordt versterkt met structuren, zoals richels, en vooral bij vleesetende soorten een aantal tanden kan hebben. De mondholte mondt uit in een gespierde, zuigende farynx, ook bekleed met de cuticula. Spijsverteringsklieren zitten in dit deel van het spijsverteringskanaal, die de enzymen voor de vertering uitscheiden. Bij soorten met een stilet kunnen deze verteringsenzymen in de prooi worden geïnjecteerd.[11] Aan het eind van de farynx zit een verdikking voor het vermalen van het voedsel.
Er is geen maag aanwezig. De farynx sluit rechtstreeks aan op een spierloos gedeelte, dat de grootste lengte van het spijsverteringskanaal omvat. Hier worden meer enzymen geproduceerd en worden de voedingsstoffen door een één cel dikke laag geabsorbeerd. Het laatste deel van het spijsverteringskanaal is het rectum dat ook bekleed is met de cuticula. Het achterste deel van het rectum tot aan de anus wordt het proctodeum genoemd. Net onder en voor de punt van de staart zit de anus. Door de lichaamsbewegingen van de rondworm wordt het voedsel door het spijsverteringkanaal voortbewogen. Het spijsverteringskanaal heeft kleppen of sluitspieren aan beide uiteinden voor het beheersen van de beweging van het voedsel door het lichaam.[11]
Bij de Trichocephalida en Mermithida is in sommige stadia een zeer opvallend stichosoom aanwezig, die bestaat uit een in de lengterichting liggende reeks van klierachtige, eencellige cellen, stichocyten, gerangschikt in een rij langs de farynx en de achterste farynxklieren vormen. Het mond uit in de farynxholte en functioneert blijkbaar als een uitscheidingsklier en opslagorgaan. De stichocyten bevatten mitochondria, ruw endoplasmatisch reticulum, veel Golgi-apparaten en meestal 1 van 2 soorten uitgescheiden granulen, α-granulen en β-granulen, die duiden op de uitscheidingsfunctie van de stychocyt.
Staart
Aan het uiteinde van de staart wordt vaak een kleverige 'staartklier' gevonden.[11]
Uitscheidingssysteem
Het te veel aan stikstof wordt in de vorm van ammoniak via de lichaamswand uitgescheiden en is niet gebonden aan specifieke organen. De structuren voor het uitscheiden van zout voor het op peil houden van de osmoregulatie zijn echter meestal complexer.[11]
Bij veel in zee levende rondwormen scheiden een of twee eencellige 'renette' klieren het zout uit in de vorm van 0,5-1,5 µm grote korrels door een porie aan de onderkant van het lichaam, even voorbij de farynx. Bij de meeste andere rondwormen zijn deze gespecialiseerde cellen vervangen door een orgaan bestaande uit twee parallelle kanaaltjes verbonden door een enkel dwars kanaaltje. Dit dwarse kanaaltje komt uit in een gemeenschappelijk kanaaltje dat uitkomt bij de uitscheidingsopening.[11] Met de uitscheidingen kunnen de rondwormen zich vastzetten op de ondergrond.[17]
Zenuwstelsel
Vier perifere zenuwen lopen over de lengte van het lichaam, dorsaal (rugzijde), ventraal (buikzijde) en lateraal (zijkant). Elke zenuw wordt omgeven door bindweefsel dat onder de huid en tussen de spiercellen ligt. De ventrale zenuw is de grootste en heeft een dubbele structuur naar de excretieopening. De dorsale zenuw is verantwoordelijk voor motorische controle, terwijl de laterale zenuwen sensorisch van aard zijn en de ventrale zenuw combineert beide functies. [11]
De enige plaats op het lichaam van de rondworm waar onbeweeglijke trilharen voorkomen, en die een sensorische functie hebben, zijn de zenuwbanen.[18][19]
Aan het voorste uiteinde van de rondworm vertakken de zenuwen zich vanuit een dichte, cirkelvormige zenuw (zenuwring) rondom de farynx en dient als een soort hersenen. Kleinere zenuwen lopen naar voren vanuit de ring voor de sensorische organen van de kop.[11]
De lichamen van de rondwormen zijn bedekt met talloze sensorische haren en papillen die samen zorgen voor een gevoel van aanraking. Achter de sensorische haren op de kop liggen twee kleine putjes, of amfiden. Dit zijn de belangrijkste reukorganen van een rondworm. Elke amfide bestaat uit 12 sensorische zenuwcellen en met trilhaartjes bezette dendrieten. Enkele in het water levende rondwormen bezitten wat pigmentvlekken, die op oogvlekken lijken, maar het is onduidelijk of deze daadwerkelijk sensorisch van aard zijn.[11]
"Spierarmen" zijn in de huid voorkomende armvormige spiermembraanverlengingen, die de in de huid voorkomende spiermembranen met de motoraxonen van de dorsale zenuwbaan en die van de ventrale zenuwbaan verbinden. In tegenstelling tot andere organismen waar neuronen prikkels naar hun doelspiercellen sturen om synapsen te maken, worden neuromusculaire knooppunten van Caenorhabditis elegans gemaakt door armvormige uitgroeiïngen (spierarmen) van de spiercellen naar de motorneuronen. Deze spierarmen bestaan uit eenvoudige structuren en hebben een steel met een vertakte terminus die contact maakt met het neuron. Vergelijkbaar met chemische synapsen tussen neuronen, worden neuromusculaire knooppunten terloops gemaakt door de gestimuleerde neuronen aan deze spierarmen.[20]
-
 Lichaamsbouw van de mannelijke organen van een Caenorhabditis elegans (In werkelijkheid is het aaltje kleurloos en doorschijnend.)
Lichaamsbouw van de mannelijke organen van een Caenorhabditis elegans (In werkelijkheid is het aaltje kleurloos en doorschijnend.) -
 Lichaamsbouw van de mannelijke organen van een Caenorhabditis elegans (In werkelijkheid is het aaltje kleurloos en doorschijnend.)
Lichaamsbouw van de mannelijke organen van een Caenorhabditis elegans (In werkelijkheid is het aaltje kleurloos en doorschijnend.) -
 Lichaamsbouw van de vrouwelijke organen van de hermafrodiete Caenorhabditis elegans (In werkelijkheid is het aaltje kleurloos en doorschijnend.)
Lichaamsbouw van de vrouwelijke organen van de hermafrodiete Caenorhabditis elegans (In werkelijkheid is het aaltje kleurloos en doorschijnend.) -

-
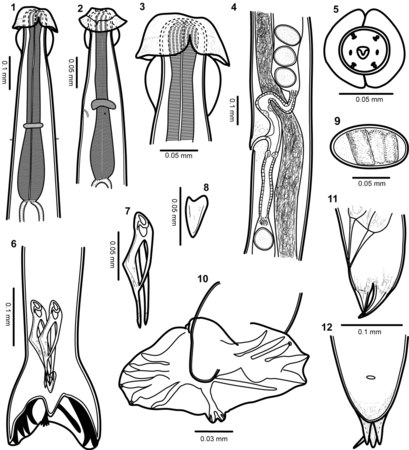 Lichaamsbouw van Torrestrongylus tetradorsalis. 1: Voorste uiteinde van mannetje, buikaanzicht, met de farynx en de relatieve posities van de zenuwring en deiriden (sensorische papillen als tastorgaan in het laterale cervicale gebied); let op scheiding van cuticulaire projectie in voorste blaasje. 2: Voorste uiteinde van vrouwtje, zijaanzicht, dat de relatieve posities van de zenuwring, deiride en uitscheidingsporie toont; let op continue cuticulaire expansie in voorste blaasje. 3: Detail van de voorste helft van het kopblaasje met de laterale uitsteeksels die een paraplu vormen en de achterste helft met de vorm van de "twee handvatten van een kruik". 4: Ovejector (het terminale, zeer gespierde deel van de eileider, die het ei door de genitale porie duwt) van vrouwtje met flap in voorste lip van vulva, vagina vera, infundibulum (tweede deel van de eileider), sluitspieren en baarmoedervertakkingen met uitgegroeide eieren. 5: Gezichtsaanzicht van het mannetje, met de rangschikking van papillen en cuticulaire vergroting van de voorste kopblaasje. 6: Achterste uiteinde van het mannetje, dat de relatieve positie van spicula en gubernaculum toont. 7: Spiculum. 8: Gubernaculum. 9: Ei. 10: Staartzak met plaats en vertakking van dorsale straal. 11: Zijaanzicht van de staart van het vrouwtje. 12: Buikaanzicht van vrouwelijke staart.
Lichaamsbouw van Torrestrongylus tetradorsalis. 1: Voorste uiteinde van mannetje, buikaanzicht, met de farynx en de relatieve posities van de zenuwring en deiriden (sensorische papillen als tastorgaan in het laterale cervicale gebied); let op scheiding van cuticulaire projectie in voorste blaasje. 2: Voorste uiteinde van vrouwtje, zijaanzicht, dat de relatieve posities van de zenuwring, deiride en uitscheidingsporie toont; let op continue cuticulaire expansie in voorste blaasje. 3: Detail van de voorste helft van het kopblaasje met de laterale uitsteeksels die een paraplu vormen en de achterste helft met de vorm van de "twee handvatten van een kruik". 4: Ovejector (het terminale, zeer gespierde deel van de eileider, die het ei door de genitale porie duwt) van vrouwtje met flap in voorste lip van vulva, vagina vera, infundibulum (tweede deel van de eileider), sluitspieren en baarmoedervertakkingen met uitgegroeide eieren. 5: Gezichtsaanzicht van het mannetje, met de rangschikking van papillen en cuticulaire vergroting van de voorste kopblaasje. 6: Achterste uiteinde van het mannetje, dat de relatieve positie van spicula en gubernaculum toont. 7: Spiculum. 8: Gubernaculum. 9: Ei. 10: Staartzak met plaats en vertakking van dorsale straal. 11: Zijaanzicht van de staart van het vrouwtje. 12: Buikaanzicht van vrouwelijke staart. -
 Lichaamsbouw van Capillaria plectropomi. A: voorste uiteinde van mannetje, zijaanzicht. B: stichocyte in het middelste deel van het stichosoom. C: achterste uiteinde van mannetje, zijaanzicht. D: kop van vrouwtje, apicaalaanzicht. E: staarteinde van mannetje, ventraalaanzicht. F: regio van vulva, zijaanzicht. G, H: staarteinde van mannetje verschillende exemplaren), zijaanzichten. I: staart van vrouwtje, zijaanzicht. J: spiculum, zijaanzicht. K: laterale bacillaire band in de farynx, zijaanzicht. L: volledig ontwikkeld ei.
Lichaamsbouw van Capillaria plectropomi. A: voorste uiteinde van mannetje, zijaanzicht. B: stichocyte in het middelste deel van het stichosoom. C: achterste uiteinde van mannetje, zijaanzicht. D: kop van vrouwtje, apicaalaanzicht. E: staarteinde van mannetje, ventraalaanzicht. F: regio van vulva, zijaanzicht. G, H: staarteinde van mannetje verschillende exemplaren), zijaanzichten. I: staart van vrouwtje, zijaanzicht. J: spiculum, zijaanzicht. K: laterale bacillaire band in de farynx, zijaanzicht. L: volledig ontwikkeld ei.






Levenscyclus

Als voorbeeld wordt die van de wortelknobbelaaltjes beschreven.
Alle rondwormen hebben een embryonaalstadium, vier larvale stadia (L1 - L4) en een volwassen stadium. Het jonge Meloidogynes-aaltje komt uit het ei als een vermiforme larve in het tweede stadium (L2), waarbij de eerste vervelling in het ei heeft plaatsgevonden. Pas uitgekomen larven hebben een korte vrijlevende fase in de bodem, in de rhizosfeer van de waardplanten. Ze kunnen de ouderwaardplanten opnieuw infecteren of door de grond migreren naar een nieuwe waardplantwortel. L2-larven voeden zich niet tijdens de vrijlevende fase, maar leven van in de darm opgeslagen lipiden.[21]
Een uitstekend modelsysteem voor de studie van het parasitaire gedrag van plantparasitaire aaltjes is ontwikkeld met behulp van de zandraket (Arabidopsis thaliana) als modelgastheer.[22] De wortels van de zandraket zijn aanvankelijk klein en transparant, zodat elk detail zichtbaar is. Binnendringen en migratie in de wortel werd bestudeerd met Meloidogyne incognita.[23] Aaltjes van het tweede larvale stadium dringen het wortelstrekkingsgebied binnen en migreren in de wortel totdat ze een vaste verblijfplaats hebben gevonden. Signalen van de L2-aaltjes zorgen ervoor dat parenchymcellen in de buurt van de kop van het L2-aaltje meerkernig worden[24] voor het vormen van voedingscellen, algemeen bekend als reuzencellen, van waaruit de L2 en later de volwassen aaltjes zich voeden met behulp van een stilet.[25] Gelijktijdig met de vorming van reuzencellen, vormt het omringende wortelweefsel een gal (knobbel) waarin de zich ontwikkelende larve zit. De larven beginnen zich met de inhoud van de reuzencellen te voeden ongeveer 24 uur nadat ze een vaste verblijfplaats hebben gevonden.
Na verdere voeding ondergaan de L2-aaltjes morfologische veranderingen en worden ze bolvormig. Zonder verdere voeding vervellen ze drie keer en worden uiteindelijk volwassen. Bij vrouwtjes, die bijna bolvormig zijn, wordt de voeding hervat en ontwikkelt het voortplantingssysteem zich.[21] De mannetjes kunnen zich vrij bewegen en verlaten de wortel nadat ze volwassen zijn geworden. De levensduur van een volwassen vrouwtje kan tot drie maanden oplopen en er kunnen honderden eieren worden geproduceerd. Vrouwtjes kunnen na de oogst van de bovengrondse delen van de plant doorgaan met het leggen van eieren. De overlevingsfase van het aaltje tussen de opeenvolgende gewassen bevindt zich meestal in het ei.
De lengte van de levenscyclus is temperatuurafhankelijk.[26][27] De relatie tussen ontwikkelingssnelheid en temperatuur is lineair over een groot deel van de levenscyclus van de wortelknobbelaaltjes, hoewel het mogelijk is dat de verschillende stadia van de levenscyclus, bijvoorbeeld ei-ontwikkeling, gastheer wortelinvasie van de waardplant of groei, een iets ander optimum hebben. Soorten binnen het geslacht Meloidogyne kunnen ook verschillende temperatuuroptima's hebben. Bij bijvoorbeeld Meloidogyne javanica vindt de ontwikkeling plaats tussen 13 en 34 °C, met een optimale ontwikkeling bij ongeveer 29 °C.
De vrouwtjes leggen de eieren in een gelatineuze massa geproduceerd door zes rectale klieren en afgescheiden voor en tijdens het leggen van de eieren.[28][29] De gelatineuze massa vormt aanvankelijk een kanaal door de buitenste lagen van het wortelweefsel en omringt later de eieren. De gelatineuze massa vormt een barrière tegen waterverlies en zorgt door het hoge vochtgehalte dat de eieren niet uitdrogen.[30] Naarmate de gelatineuze massa ouder wordt, verandert het van een kleverige, kleurloze gelei in een leerachtige, oranje-bruine substantie die gelaagd lijkt.[31]
De eivorming is in Meloidogyne javanica gedetailleerd bestudeerd[32] en lijkt op de eivorming in het goed bestudeerde, vrij levende aaltje Caenorhabditis elegans.[33] Ook de embryogenese is bestudeerd en de ontwikkelingsstadia in een geplette eimassa zijn gemakkelijk te identificeren met een fasecontrastmicroscoop. Het ei bestaat uit één cel, met herkenbare volgende stadia van twee cellen, vier cellen en acht cellen. Verdere celdeling leidt tot het kikkervisjesstadium, waarbij door een verdere verlenging het eerste larvale stadium ontstaat. Het aaltje is nu ruwweg vier keer zo lang als het ei was. Het L1-stadium van Caenorhabditis elegans heeft 558 cellen en het L1-stadium van Meloidogyne javanica heeft waarschijnlijk een vergelijkbaar aantal, omdat alle aaltjes morfologisch en anatomisch vergelijkbaar zijn.[33] Het omhulsel van het ei bestaat uit drie lagen, met als buitenste laag de vitellinelaag, dan een chitine-achtige middelste laag en een binnenste laag bestaande uit lipiden. De vitellinelaag bestaat voornamelijk uit eiwitvezels, met eiwitreceptoren die nodig zijn voor de spermabinding die op hun beurt gebonden zijn aan spermaplasmamembraanreceptoren.
Nadat het eiomhulsel doorlaatbaar is geworden, kan de larve uit het ei komen met behulp van fysieke en/of enzymatische processen.[34] Eitjes van de wortelknobbelaaltjes worden in het algemeen niet beïnvloed door de aanwezigheid van een gastheer, maar komen pas uit bij de juiste temperatuur en in aanwezigheid van water. Ammonium-ionen kunnen het uitkomen remmen en het binnendringingsvermogen in de plantenwortels door de Meloidogyne incognita -larven verminderen. [35]
Wijze van voortplanting
Rondwormen kunnen eenslachtig of hermafrodiet zijn. De meeste rondwormen zijn eenslachtig met zowel vrouwelijke als mannelijke exemplaren. Bij sommige soorten zoals Caenorhabditis elegans zijn de meeste rondwormen hermafrodiet en komen er slechts enkele mannelijke exemplaren voor. Rondwormsoorten vertonen een reeks van verschillende wijzen van voortplanting, waaronder seksuele voortplanting (amfimixis), facultatieve seksuele voortplanting, meiotische parthenogenese (automixie) en mitotische parthenogenese (apomixie).[36]
Beide geslachten bezitten een of twee buisvormige gonaden. Bij de mannetjes wordt het sperma geproduceerd aan het einde van de geslachtsklieren en verplaatst het zich tijdens de rijping over de lengte van de rondworm. De testis komt uit in een relatief brede zaadblaas en gaat vervolgens over in een beklierd en gespierd ejaculatiekanaal dat te vergelijken is met de zaadleider en cloaca. Bij de vrouwtjes komen de twee eierstokken uit in twee eileiders (bij hermafrodieten komen de eieren eerst in een spermatheca) en vervolgens in een beklierde baarmoeder. De baarmoeders komen beide uit in een gemeenschappelijke vulva/vagina, meestal gelegen aan de buitenkant in het midden van de morfologisch buikzijde.[11]
De voortplanting is meestal seksueel, hoewel hermafrodieten in staat zijn tot zelfbevruchting. Mannetjes zijn meestal kleiner (vaak veel kleiner) dan vrouwtjes of hermafrodieten en hebben vaak een karakteristiek gebogen of waaiervormige staart. Aan de staartzijde van de mannelijke rondworm kunnen 1 of 2 spicula voorkomen, die dienen om de vulva van de vrouwelijke rondwormen te openen en de overdracht van sperma te vergemakkelijken, hoewel sperma niet rechtstreeks door of met de spicula wordt overgedragen.[13][37] Het gubernaculum is een ander orgaan van het mannelijk copulatiesysteem, dat de spiculum of de spicula tijdens de copulatie geleidt. Tijdens de copulatie kruipen amoebeachtig zaadcellen langs het spiculum in de vulva van het vrouwtje. Aangenomen wordt dat de zaadcel van rondwormen de enige eukaryotische cel is zonder het bolvormige eiwit G-actine.
Eieren kunnen een embryo of nog geen embryo bevatten op het moment dat ze door het vrouwtje worden uitgescheiden. Er is nog geen embryo aanwezig als de bevruchte eieren nog niet zijn ontwikkeld. Van een paar soorten is bekend dat ze eierlevendbarend zijn. Het omhulsel van het ei bestaat uit drie lagen, met als buitenste laag de vitellinelaag, dan een chitine-achtige middelste laag en een binnenste laag bestaande uit lipiden. De vitellinelaag bestaat voornamelijk uit eiwitvezels, met eiwitreceptoren die nodig zijn voor de spermabinding die op hun beurt gebonden zijn aan spermaplasmamembraanreceptoren. Bij vrij levende rondwormen vinden de eerste en tweede larvale stadia al in het ei plaats. Bij parasitaire rondwormen is de levenscyclus vaak veel gecompliceerder.[11] Nadat het eiomhulsel doorlaatbaar is geworden, kan het embryo of de larve uit het ei komen met behulp van fysieke en/of enzymatische processen.[34]
Bijzondere vormen van voortplanting
Sommige hermafrodiete rondwormen, zoals Heterorhabditis-soorten van de orde Rhabditida, hebben een wijze van voortplanting, genaamd endotokia matricida (in de uterus geboren), wat de dood van de moeder tot gevolg heeft. Dit komt vooral voor bij entomopathogene rondwormen, rondwormen die van insecten leven. De larven eten de moeder van binnen uit op en laten alleen het exoskelet achter. Deze wijze van voortplanting verzekert de ontwikkeling van de larven op een moment dat de externe voedselvoorziening afneemt en zorgt ervoor dat de nakomelingen voldoende energiereserves en symbiotische bacteriën hebben, waardoor ze voor lange tijd zonder voedsel kunnen overleven. [38][39]
Caenorhabditis elegans en Caenorhabditis briggsae hebben androdioecie als voortplantingmethode, wat bij dieren zeer zeldzaam is. Hierbij komen zowel mannetjes als hermafrodieten tegelijk voor. De hermafrodieten geven echter bij de voortplanting de voorkeur aan de mannetjes in plaats van aan zelfbevruchting te doen. Ook geeft het paren met mannetjes meer nageslacht, doordat er dan meer eieren gelegd worden.
Rondwormsoorten van het geslacht Meloidogyne (wortelknobbelaaltjes) hebben een aantal verschillende voortplantingsmethoden, waaronder seksuele voortplanting (amfimixis), facultatieve seksuele voortplanting, meiotische parthenogenese (automixie) en mitotische parthenogenese (apomixie).[36]
Het geslacht Mesorhabditis vertoont een ongebruikelijke vorm van parthenogenese, waarbij sperma producerende mannetjes copuleren met vrouwtjes, maar de spermacel niet fuseert met de eicel. Contact met de spermacel is essentieel voor de eicel om met delen te beginnen, maar omdat er geen fusie van de cellen optreedt, draagt het mannetje geen genetisch materiaal over aan de nakomelingen, die in wezen klonen van het vrouwtje zijn.[11]
Taxonomie
Over de systematiek van de rondwormen (nematoden) bestaat geen consensus.
Pathotypen
Bij soorten van plantenparsitaire aaltjes kunnen pathotypen voorkomen. Een pathotype is een systematische eenheid binnen een aaltjessoort, die niet is te onderscheiden op morfologische kenmerken. Pathotypen verschillen van elkaar doordat ze bepaalde resistenties bij plantenrassen van één of enkele waardplantsoorten hebben doorbroken. Zo zijn van Globodera rostochiensis (het aardappelcystenaaltje) momenteel 5 pathotypen bekend: Ro1, Ro2, Ro3, Ro4 en Ro5. Ro1 komt in Nederland het meeste voor en Ro4 en Ro5 het minste. Zo heeft het wortelknobbelaaltje Meloidogyne arenaria 3 pathotypen, waarvan pathotype 1 het meest voorkomt in Amerika op pinda.
Vrij levende soorten
Vrij levende soorten voeden zich met algen, schimmels, kleine dieren, uitwerpselen, dode organismen en levend weefsel. Vrij levende mariene rondwormen zijn belangrijk en komen veel voor bij de meiobenthos. Ze spelen een belangrijke rol in het afbraakproces en helpen bij het opnieuw beschikbaar komen van voedingsstoffen in mariene omgevingen. Ze zijn echter gevoelig voor veranderingen in het milieu veroorzaakt door vervuiling. Caenorhabditis elegans is een in de bodem vrijlevende rondworm, die is gebruikt als modelorganisme. Onder de vrijlevende rondwormen komen plantenparasitaire soorten voor, zoals bladaaltjes, stengelaaltjes en wortellesieaaltjes. Van de wortelknobbelaaltjes zijn de volwassen mannetjes ook vrijlevend.
Parasitaire soorten


Parasitaire soorten rondwormen, die bij mensen voorkomen, zijn onder andere de Ascaris-soorten, zoals de spoelworm, Filarioidea, Ancylostoma, Necator, aarsmade en zweepworm (Trichuris trichiura). Trichinella spiralis komt voor bij ratten, varkens, beren en bij mensen dor het eten van besmet vlees. Veroorzaakt de ziekte trichinose of trichinellose. Baylisascaris infecteert meestal wilde dieren, maar kan voor mensen ook dodelijk zijn. De hartworm (Dirofilaria immitis) infecteert de harten, slagaders en longen van honden en sommige katten. De rode lebmaagworm (Haemonchus contortus) is een van de meest voorkomende besmettelijke ziekten over de hele wereld bij schapen.
Entomopathogene nematoden parasiteren daarentegen op schadelijke insecten en worden door de mens meestal als gunstig beschouwd, maar ze infecteren in sommige gevallen ook nuttige insecten. Ze dringen de insecten binnen en de bacteriën, die als symbiont in hun lichaam leven, doden vervolgens het insect. De belangrijkste hiervan, die gebruikt worden bij de biologische bestrijding van schadelijke insecten, behoren tot de geslachten Heterorhabditis en Steinernema.
Een soort rondworm parasiteert alleen de gewone vijgenwesp, terwijl deze wesp het enige insect is die zorgt voor de bevruchting van de vijgen. De bevruchting van de vijgenboom geschiedt namelijk door symbiose met de gewone vijgenwesp, die zowel zichzelf als de vijg voortplant. Een bevruchte vrouwelijke, geparasiteerde wesp gaat de vijg door een opening (ostiole) binnen, bevrucht de vrouwelijke vijgenbloemen met stuifmeel en legt haar eieren in deze bloemen, waarna ze sterft. De nakomelingen van de rondworm gaan met de volgende generatie wespen mee naar nieuwe vijgen.
_%26_Mermithid_nematode_(black_background).JPG)
De nieuw ontdekte parasitaire Tetradonematidae-rondworm Myrmeconema neotropicum induceert fruitmimicry bij de tropische mier Cephalotes atratus. Geïnfecteerde mieren ontwikkelen een felrood achterlijf, hebben de neiging om zich trager te bewegen en lopen met hun achterlijf in een opvallend omhoog gerichte positie. Deze veranderingen trekken waarschijnlijk bes etende vogels aan, die de geïnfecteerde mieren opeten. De parasieteneieren komen in de uitwerpselen van de vogels terecht, die door de mieren worden verzameld om als voedsel voor hun larven te dienen, waarmee de levenscyclus van de rondworm M. neotropicum is voltooid.
Evenzo zijn meerdere soorten rondwormen gevonden in de buikholten van de zweetbij, Lasioglossum zephyrum. In het vrouwelijk lichaam belemmert de rondworm de ontwikkeling van de eierstokken en maakt de bij minder actief, dus minder effectief bij het verzamelen van stuifmeel. [40] Ook wordt een Mermithidae-rondworm aangetroffen bij de Aziatische hoornaar.
Bij insecten behorend tot de superfamilie Apoidea werden de volgende rondwormen gevonden. De Dufour's klieren van de vrouwtjes van Halictus ligatus, Augochlora pura mosieri en Augochlorella gratiosa (Halictidae) uit Florida waren geïnfecteerd met de rondwormlarven van Aduncospiculum halicti (Diplogasteridae). De Dufour's klieren van Colletes thoracicus (Colletidae) vrouwtjes uit Maryland waren geïnfecteerd met de larven van een nieuw soort rondworm behorend tot Koerneria (Diplogasteridae) en de klieren in de buikholte van de vrouwelijke Andrena alleghaniensis (Andrenidae) uit New York waren geïnfecteerd met de larven van een andere Koerneria-soort. De eileiders aan de buikzijde en op het mediane vlak, de Dufour's klieren en de gifklieren van Anthophora abrupta (Anthophoridae) uit Maryland en Alabama waren geïnfecteerd met de larven van een nieuwe rondwormsoort behorend tot Bursaphelenchus (Aphelenchoididae).[41]
Land-, tuin- en bosbouw
Afhankelijk van de soort kan een rondworm nuttig of schadelijk zijn voor planten.
Insectenparasitaire rondwormen kunnen schadelijke insecten bestrijden, zoals de inzet van Steinernema feltiae bij de bestrijding van de eikenprocessierups[2], het gebruik van Phasmarhabditis hermaphrodita tegen naaktslakken en bij mais de rondworm Steinernema carpocapsae bij de bestrijding van Helicoverpa zea.[42]
Plantenparasitaire aaltjes omvatten verschillende groepen, die ernstige gewasverliezen veroorzaken. De meest voorkomende geslachten met plantenparasitaire aaltjessoorten zijn Aphelenchoides (bladaaltjes), Ditylenchus, Globodera (aardappelcystenaaltjes), Heterodera, Longidorus, Meloidogyne (wortelknobbelaaltjes), Nacobbus, Pratylenchus (wortellesieaaltjes), Trichodorus en Xiphinema. Verschillende plantenparasitaire aaltjes veroorzaken schade aan de wortelweefsels, inclusief de vorming van zichtbare gallen, zoals de wortelknobbelaaltjes.
Rondwormen kunnen schadelijke virussen overdragen, zoals Xiphinema spp, Longidorus spp, Trichodorus spp. Xiphinema diversicaudatum kan het arabis-mozaïekvirus over brengen bij Fraxinus en het strawberry latent ringspot virus bij aardbeien. X. americanum is een overbrenger van het tabaksringvlekvirus (tobacco ringspot virus).[43] Xiphinema index draagt het grapevine fanleaf virus over.
Andere rondwormen tasten bomen aan. De belangrijkste vertegenwoordiger van deze groep is Bursaphelenchus xylophilus, de rondworm dat in dennenbossen voorkomt in Azië en Amerika en recent ontdekt is in Europa.
Parasitaire plantenaaltjes kunnen bestreden worden door vruchtwisseling of gebruik te maken van resistente rassen. Bij het gebruik van resistente rassen kunnen echter pathotypen gaan optreden, zoals bij het aardappelcystenaaltje, waarvan thans 5 pathotypen bestaan. Het blijft dan een race tussen de plantenverdelaar en de aanpassingssnelheid van het aaltje. Ook kunnen plantensoorten als Tagetes patula of andere Tagetes-soorten ingezet worden tegen de wortellesieaaltjes.[44] Het wortelexudaat van de Tagetesplanten thiophene a-terthienyl doodt de aaltjes.[45] Een nadeel hiervan is dat er in het betreffende seizoen geen gewas geteeld kan worden. Een andere mogelijkheid is het gebruik van natuurlijke antagonisten zoals de schimmel Gliocladium roseum, waarvan de conidia zich hechten aan de cuticula van het aaltje, ontkiemen en met hun kiemhyfen het aaltje binnendringen en doden.[46] Chitosan, een natuurlijk biocide, lokt plantenafweerreacties uit waarbij parasitaire cystenaaltjes op de wortels van soja, maïs, suikerbiet, aardappel en tomaat gedood worden, maar niet de niet-parasitaire aaltjes in de bodem.[47] Stomen van de grond, waarbij de grondtemepratuur renminste moet oplopen tot 80°C, is een effectieve methode om aaltjes te doden voordat een gewas wordt geplant, maar doodt zowel de schadelijke als de nuttige bodemfauna. CSIRO heeft een 13- tot 14-voudige afname van de populatiedichtheden van Meloidogyne javanica-aaltjes gevonden in percelen met het sareptamosterdras Nemfix (Brassica juncea var. juncea) als groenbemester, dat dient als vanggewas voor de aaltjes.[48] Door de wortels van het vanggewas worden de larven uit de cysten gelokt, maar ze kunnen zich hierop niet vermeerderen. Ook kan bladrammenas als groenbemester ingezet worden tegen het bietencystenaaltje en aardappelmoeheid, maar bladrammenas is een waardplant van het witte (Heterodera schachtii) en het gele bietencystenaaltje (Heterodera trifolii). Daarom moeten er tegen deze aaltjes resistente rassen van bladrammenas worden gebruikt.
Parasitaire plantenaaltjes kunnen ook bestreden worden door aaltjesetende aaltjes.
Varia
In juli 2018 beweerden Russische wetenschappers tijdens bodemonderzoek in de Pleistocene Parken twee rondwormen aangetroffen te hebben in het Siberische permafrost. De dieren zouden tot 41.700 jaar oud zijn. De rondwormen werden meegenomen naar een laboratorium, waar ze langzaam aan ontdooid werden en tegen alle verwachtingen in terug tot leven kwamen en opnieuw volledig functioneerden. Hiermee zouden deze rondwormen de langst levende dieren ooit zijn op onze planeet. Terwijl er in de wetenschappelijke wereld nog twijfels bestaan over de juistheid van deze bevindingen, wordt er algemeen aangenomen dat het theoretisch niet onmogelijk is dat deze dieren na dergelijk lange periode terug tot leven gewekt kunnen worden.[49]
Fotogalerij
-
 Meloidogyne incognita-aaltje dat een tomatenwortel binnendringt, vergroting 500X
Meloidogyne incognita-aaltje dat een tomatenwortel binnendringt, vergroting 500X -
 Volwassen Caenorhabditis elegans
Volwassen Caenorhabditis elegans -
 Een vrouwelijke Nippostrongylus brasiliensis-aaltje
Een vrouwelijke Nippostrongylus brasiliensis-aaltje -
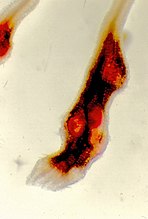 Vrouwtje van Meloidogyne arenaria in een pindawortel
Vrouwtje van Meloidogyne arenaria in een pindawortel -
 Eitjes uit opengebroken cyst van een Heteroderidae-aaltje op Rubus spp.
Eitjes uit opengebroken cyst van een Heteroderidae-aaltje op Rubus spp. -
 Eitje van een Heteroderidae-aaltje
Eitje van een Heteroderidae-aaltje -
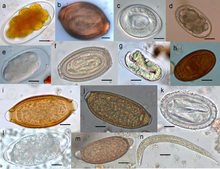 Rondwormparasieteneitjes en een Strongyloides stercoralis aaltje
Rondwormparasieteneitjes en een Strongyloides stercoralis aaltje -
 Meloidogyne arenaria L2-aalyjes in wortelpunt van de pinda
Meloidogyne arenaria L2-aalyjes in wortelpunt van de pinda -
 L3-aaltje van Phasmarhabditis hermaphrodita
L3-aaltje van Phasmarhabditis hermaphrodita -
 Striae-patroon rond de anus (perinaalpatroon) van een mannelijk Meloidogyne incognita-aaltje
Striae-patroon rond de anus (perinaalpatroon) van een mannelijk Meloidogyne incognita-aaltje -
 Levenscyclus van Strongyloides stercoralis
Levenscyclus van Strongyloides stercoralis -
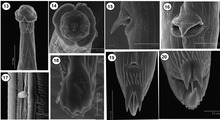
Torrestrongylus tetradorsalis. Rasterelektronenmicroscopische afbeeldingen.
-
 Kop van Philometra fasciati met papillen
Kop van Philometra fasciati met papillen -

-
 Philometra protonibeae, links vrouwtje, rechts mannetje met spiculum
Philometra protonibeae, links vrouwtje, rechts mannetje met spiculum
_penetrates_a_tomato_root_-_USDA-ARS.jpg)



.jpg)


.jpg)

.jpg)


.png)
.png)
- Onderschrift afbeelding van Torrestrongylus tetradorsalis rasterelektronenmicroscopische afbeeldingen: 13: Subventraal aanzicht van het voorste uiteinde, met laterale afbeeldingen van cefalisch (kop) blaasje; het achterste deel van het cefalisch blaasje lijkt "leeggelopen". 14: Frontaalaanzicht met mondopening, papillen en amfiden. 15: Detail van de vulva met opvallende flap, zijaanzicht. 16: Ventrale weergave van vulva met de basis van de vulvaire flap. 17: Deiride (een sensorische papil als tastorgaan in het laterale cervicale gebied). 18: Zijaanzicht van caudale bursa (staartzakje), afbeelding met bursale stralen. 19: Ventrale weergave van de staart van het vrouwtje, met anus en terminale stekels. Striae zijn zichtbaar. 20: Subterminaal beeld van de staart van het vrouwtje, met alle drie de cuticulaire afbeeldingen en terminaal filament (draadje). Striae zijn zichtbaar.
Externe links
- Plant and insect parasitic nematodes determination
- NeMys: Website over mariene nematoden
- ITIS = Integrated Taxonomy Information System
- Aaltjesschema van PPO / Universiteit Wageningen
- ProGemüse - Duits-Nederlands EU-project rondom schadelijke rondwormen in de groenteteelt
|