
Embryophyta
| Embryophyta Fossiel voorkomen: Ordovicium[1] – heden | |||
|---|---|---|---|

| |||
| Diversiteit van Embryophyta | |||
| Taxonomische indeling | |||
| |||
| Onderrijk | |||
| Embryophyta Engl. (1892) | |||
| Afbeeldingen op | |||
| Embryophyta op | |||
| |||
Embryophyta is een botanische naam voor een groep planten waarbij de individuele planten uitgroeien vanuit een embryo. Het gaat om de landplanten, dus ruwweg de mossen, varens en zaadplanten.
De naam landplanten is ter onderscheid met de algen. Onder de landplanten bevinden zich ook verschillende planten van het zoete en enkele van het zoute water. Een andere naam voor een vergelijkbare groep is Cormophyta. Deze naam staat dan tegenover Thallophyta, voor 'planten' die geen differentiatie in stengel, blad en wortel vertonen. De mossen vormen bij deze indeling een groep dicht bij de grens tussen de twee groepen; men spreekt bijvoorbeeld van thalleuze en folieuze levermossen, waarmee wordt gedoeld op de gametofyt en niet op de sporofyt.
Alle soorten in deze taxonomische groep zijn complexe meercellige eukaryoten met gespecialiseerde voortplantingsorganen. Met slechts een beperkt aantal uitzonderingen verkrijgen ze hun energie door fotosynthese.
Flora's[bewerken | brontekst bewerken]
De 23e druk van de Heukels' Flora van Nederland behandelt alleen de in Nederland inheemse vaatplanten, met een nadruk op de zaadplanten, de Spermatopsida. De Heukels is een flora, dus een lijst van in Nederland voorkomende planten, en gaat niet in op de vraag hoe deze groepen geplaatst moeten worden wat betreft de taxonomie in hogere rangen. Wel verwijst deze flora expliciet naar de webpagina van Tolweb.[2]
De Beknopte mosflora van Nederland en België[3] behandelt de mossen van de in Nederland en België inheemse mossen.
Geslachtelijke voortplanting[bewerken | brontekst bewerken]
De geslachtelijke voortplanting bij Embryophyta (landplanten) is een gecompliceerd proces, dat goed begrepen kan worden door vergelijking van het eenvoudiger proces bij mossen, levermossen, hauwmossen en varens met het gecompliceerde proces bij naaktzadigen en bij bedektzadigen. De overeenkomsten in de generatiewisseling werden reeds in 1849 beschreven door Wilhelm Hofmeister. Er is een grote verscheidenheid in de levenscyclus, waarvan de geslachtelijke voortplanting deel van uitmaakt, maar er zijn enkele belangrijke punten van overeenkomst.
De Embryophyta hebben als groep het land veroverd. Ze zijn daar de meest dominante groep van organismen, en zorgen voor verreweg het grootste aandeel in de primaire productie van de biomassa.
De Embryophyta hebben in de evolutie hun afhankelijkheid van water bij de voortplanting verloren. Hoewel de hele groep van de Embryophyta "landplanten" wordt genoemd, zijn er veel soorten die secundair zijn aangepast aan het leven in het zoete, brakke of zoute water. De naam "landplanten" is vooral bedoeld als tegenstelling tot de vooral waterbewonende algen, die oorspronkelijk ook wel tot de "planten" werden gerekend.
Sporen hadden oorspronkelijk een belangrijke rol bij de ongeslachtelijke voortplanting, maar met het ontstaan van de geslachtsverdeling met (mannelijke) microsporen en (vrouwelijke) macrosporen is deze rol in de loop van de fylogenie verloren gegaan. Ook de geslachtsorganen (antheridium en archegonium) zijn in de evolutie opgenomen in de omhullende beschermende structuren, zodat er meer gecompliceerde geslachtsorganen zijn ontstaan, zoals meeldraden, stampers en zaadknoppen. De antheridia en archegonia zijn microscopisch klein en nog met moeite te herkennen.[4]
Verloop van de levenscyclus[bewerken | brontekst bewerken]
Wilhelm Hofmeister onderkende omstreeks 1849 als eerste de overeenkomst in de generatiewisseling van mossen (en verwanten), varens (en verwanten) en zaadplanten, met name de coniferen, en de homologie van de organen voor de voortplanting bij verschillende groepen van de Embryophyta (landplanten).
De Embryophyta hebben alle een levenscyclus met een afwisseling van twee generaties: met een haploïde generatie, de gametofyt (de gametenvormende generatie), en een diploïde generatie, de sporofyt (de sporenvormende generatie). Een generatie is een stadium in de ontwikkeling van een organisme, dat begint met een voortplantingscel (spore of zygote), en die - na een periode van duidelijke vegetatieve activiteit zoals groei en ontwikkeling - eindigt met de vorming van andere reproductieve cellen (sporen of gameten). Een levenscyclus met een dergelijke afwisseling van generaties heet een digenetische cyclus.
Bij de Embryophyta begint de haplofase (haploïde kernfase) of gametofyt-fase met de door meiose of reductiedeling gevormde eencellige sporen, die uitgroeien tot gametofyt (de generatie die de haploïde gameten: eicellen, zaadcellen vormt). De diplofase (diploïde kernfase) of sporofytfase begint met de diploïde zygote (de bevruchte eicel), die zich via het embryo-stadium ontwikkelt tot een meercellige sporofyt (de generatie die de sporen vormt).
De belangrijkste processen in de levenscyclus zijn de bevruchting en de meiose. Hiertussen vinden de groei en ontwikkeling plaats tot een meercellig individuen: de gametofyt en de sporofyt die elkaar in een cyclus afwisselen, de digenetische cyclus.
| Schema van de levenscyclus bij Embryophyta | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
In het algemeen zijn er in de levenscyclus van Embryophyta de volgende processen en de stadia van groei en ontwikkeling te onderscheiden:
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Heterospore planten met tweehuizige sporofyten[6] | |
|---|---|
|
|
Het bovenstaande laat het principe van generatiewisseling met kernfasewisseling zien:
- de gametofyt (de gametenvormende generatie) wisselt af met de sporofyt (de sporenvormende generatie); dit type generatiewisseling is de digenetische cyclus.
- de haploïde generatie (haplofase) wisselt hier af met een diploïde generatie (diplofase); een organisme met een dergelijke kernfasewisseling is een diplohaplont.
De reductiedeling of meiose vindt plaats bij de vorming van de sporen; dit heet "sporische meiose".
Er bestaat een grote verscheidenheid in de onderlinge verhoudingen tussen sporofyt en gametofyt. De "mosplant" is de bij mossen dominante gametofyt. Na de bevruchting ontwikkelt de sporofyt zich op de gametofyt en is daarvan afhankelijk. Bij de vaatplanten (varens en zaadplanten) is de "plant" de dominante sporofyt, die vrijwel meteen uitgroeit tot de zelfstandige plant. "Dominant" wil hier zeggen: van beide generaties het sterkst ontwikkeld en het langst levend. Bij de zaadplanten is de mannelijke, maar vooral de vrouwelijke gametofyt sterk gereduceerd en vaak nauwelijks herkenbaar.
De sporofyt heeft bij mossen en varens een functie bij de ongeslachtelijke voortplanting doordat door meiotische delingen de haploïde (n) sporen in grote aantallen gevormd worden in sporangia. Bij de zaadplanten kunnen grote aantallen microsporen gevormd worden (het stuifmeel), maar in het macrosporangium (nucellus) wordt gewoonlijk slechts één functionele macrospore gevormd. De voortplanting wordt verzorgd door de grote aantallen zaadknoppen (die elk een nucellus bevatten), die bij bevruchting de zaden kunnen vormen. Hier spreekt men van geslachtelijke voortplanting door middel van zaden.
Mossen en varens[bewerken | brontekst bewerken]
Bij mossen en varens is er een sterke overeenkomst in voortplantingsorganen, hoewel hun levenscyclus sterk lijkt te verschillen. Bij varens is de variatie groot. Het eenvoudiger proces van de bevruchting bij mossen en varens wordt eerst beschreven om daarna de bevruchting bij zaadplanten te verduidelijken.
Mossen, levermossen en hauwmossen[bewerken | brontekst bewerken]
Mossen, levermossen en hauwmossen hebben een generatiewisseling met dominante haploïde gametofyt, en leeft de diploïde sporofyt (het sporenkapsel) op de gametofyt.
Sporen worden gevormd door meiose van de sporemoedercellen (sporocyten) in het sporenkapsel. De sporen zijn gewoonlijk alle van gelijke grootte (isosporie), maar heterosporie komt bij slechts enkele groepen van mossen voor. De kleinere, mannelijke sporen noemt men dan microsporen, de grotere, vrouwelijke sporen noemt men macrosporen.
De haploïde sporen groeien uit via een protonema-stadium tot een gametofyt, de "mosplant". De mosplanten vormen de geslachtsorganen (gametangia); de mannelijke gametangia zijn de antheridia en de vrouwelijke geslachtsorganen zijn de archegonia. De antheridia en archegonia hebben een beschermende wand van steriele cellen.
Het antheridium is een in het thallus verzonken of zittend tot kort gesteeld, bol tot eivormig orgaantje. In de buik van het antheridium worden de zaadcellen (antherozoïden, spermatozoïden) gevormd, die met behulp van twee flagellen kunnen zwemmen.
Het archegonium is min of meer bol, gewoonlijk flesvormig, met een buik en een hals en met een wand van steriele cellen. De eicel in de buik van het archegonium wordt bevrucht door een zaadcel uit een antheridium. Deze zaadcel heeft aan een dun vliesje water reeds genoeg om naar het archegonium te zwemmen. De cellen in het halskanaal lossen op en laten de zaadcel door.
Uit de diploïde zygote, de bevruchte eicel, ontwikkelt zich eerst een embryo en daaruit groeit de sporofyt, die parasiteert op de mosplant (de gametofyt). De sporofyt bestaat uit een voet, een seta en een sporenkapsel (sporangium of sporogoon). Bij levermossen heeft het sporenkapsel een eenvoudige bouw en heeft geen bladgroen. De sporofyt bij de hauwmossen is ongesteeld. Bij veel mossen kan de bouw van de sporofyt zeer complex zijn met chloroplasten in de cellen, huidmondjes en een centrale streng die lijkt op een vaatbundel.
- Antheridium, archegonium en jonge sporofyt bij enkele levermossen
-
 Marchantia polymorpha: antheridium en antherozoïden
Marchantia polymorpha: antheridium en antherozoïden -
 Marchantia polymorpha: archegonium
Marchantia polymorpha: archegonium -
 Porella: sporofyt overlangse doorsnede, omhuld door archegoniumrest
Porella: sporofyt overlangse doorsnede, omhuld door archegoniumrest -
 Klauwtjesmos (Hypnum)
Klauwtjesmos (Hypnum)



_9416.JPG)
Varens en varenachtigen[bewerken | brontekst bewerken]
Bij varens is er een afwisseling is van twee morfologisch verschillend gebouwde generaties, waarbij de sporofyt dominant is. De korter levende gametofyt is minder goed ontwikkeld, maar kan bij veel soorten zelfstandig leven. Dit type levenscyclus wordt een heteromorfe, digenetische cyclus met dominante sporofyt genoemd.
Veel varens zijn homospoor en eenhuizig, maar er zijn enkele groepen die heterospoor zijn: de kleinere microsporen worden gevormd in microsporangia en de grotere macrosporen worden gevormd in macrosporangia. Microsporangia en macrosporangia worden dan gevonden aan dezelfde, eenhuizige plant.
De door meiose gevormde sporen ontwikkelen zich tot de voorkiemen: het microprothallium, respectievelijk het macroprothallium, maar bij isospore varens is er geen onderscheid. De voorkiem is bij een aantal varensoorten vrijlevend, maar ontwikkelt zich bij sommige andere varensoorten zelfs binnen de beschermende sporenwand.
De zaadcellen, die flagellen hebben, hebben water nodig om naar de archegonia te zwemmen voor de bevruchting van de eicel.
Na de bevruchting ontwikkelt de zygote zich, via een embryostadium waar al de aanleg van een echte wortel herkenbaar is, tot een op zichzelf staande sporofyt, de varenplant, terwijl de prothallium afsterft. De sporofyt is de diploïde "varenplant". Deze is aanzienlijk robuuster dan het prothallium.
- Voortplantingsorganen bij varens en varenachtigen
-
 Gametofyt bij varens:
Gametofyt bij varens:
A-gametofyten, B-archegonia,
C-rizoïden, D-antheridia. -
 Selaginella: microspore met microprothallium (v=vegetatieve cel), antheridium (p, c=antheridiumwand) en spermatozoïden (a).
Selaginella: microspore met microprothallium (v=vegetatieve cel), antheridium (p, c=antheridiumwand) en spermatozoïden (a). -
 Selaginella: macrospore met rizoïden en binnen de sporewand (spm) een macroprothallium (pr) met onbevrucht archegonium (ar) en met 2 embryo's (emb1 en emb2)
Selaginella: macrospore met rizoïden en binnen de sporewand (spm) een macroprothallium (pr) met onbevrucht archegonium (ar) en met 2 embryo's (emb1 en emb2) -
 Mannetjesvaren (Dryopteris filix-mas)
Mannetjesvaren (Dryopteris filix-mas)




Zaadplanten, Spermatophyta[bewerken | brontekst bewerken]
De zaadplanten omvatten globaal twee grote groepen: de naaktzadigen (Gymnospermae) en de bedektzadigen (Angiospermae) met daarin de bloemplanten.
Alle zaadplanten zijn heterospoor en heterosporangiaat: de microsporen en macrosporen worden gevormd in van elkaar verschillende sporangia: in het microsporangium, resp. in het macrosporangium.
- microsporangia en meeldraden
-
 Pinus (den):
Pinus (den):
mannelijke strobilus met groot aantal microsporofyllen en microsporangia. -
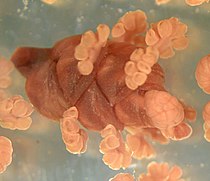 Ephedra (zeedruif):
Ephedra (zeedruif):
mannelijke strobilus met microsporofyllen = meeldraden. -
 Bedektzadigen meeldraad:
Bedektzadigen meeldraad:
A dwarsdoorsnede met 4 helmhokjes met bij 1 microsporemoedercellen, bij 2 vorming van microsporen en bij 3 rijpe stuifmeelkorrels; k=helmbindsel; s=naad waarlangs het helmhokje openbarst. -





De microsporangia van de zaadplanten groeien aan een structuur die men aanduidt met de term microsporofyl of meeldraad. Deze structuren zijn gevarieerd van vorm: bladachtig of schubachtig of asjes (al of niet vertakte stengeltjes). De microgametofyt ontwikkelt zich binnen de wand van de microspore en vormt daar een tot een of enkele cellen gereduceerd microprothallium met antheridium; gewoonlijk spreek men nu van pollen.
In het macrosporangium of nucellus worden door macrosporemoedercellen (ook wel: embryozakmoedercellen) de macrosporen gevormd. De nucellus is omgeven door een of twee beschermende integumenten. Samen vormen de nucellus met de integumenten de zaadknop. De integumenten laten aan de top van de nucellus een kleine opening, de micropyle vrij, waardoor eventueel de spermatozoïde naar de eicel kan zwemmen.
- zaadschub met zaadknop,zaadknop en vruchtbeginsel met zaadknop
-
 Pinus zaadknop overlangse doorsnede (0.6mm) A=eicel, B=archegonium, C=gametofyt, D=micropyle, E=integument
Pinus zaadknop overlangse doorsnede (0.6mm) A=eicel, B=archegonium, C=gametofyt, D=micropyle, E=integument -
 Zaadknop bij zaadplanten
Zaadknop bij zaadplanten -




Een of enkele macrosporemoedercellen ondergaan een reductiedeling of meiose, waarbij elk vier op één lijn liggende, haploïde macrosporen worden gevormd. Van deze vier macrosporen gaan in het algemeen 3 te gronde en blijft de macrospore behouden die het dichtst bij de micropile ligt: de embryozak.
Bij de naaktzadigen staan de zaadknoppen op vertakte blad- of stengelachtige structuren of op schubben (ook wel: kegelschubben), maar bij de bedektzadigen staan een of meerdere zaadknoppen op een carpel of vruchtblad of worden er door omgeven. De carpellen worden gehomologiseerd met macrosporofyllen.
Naaktzadigen, Gymnospermae[bewerken | brontekst bewerken]
De groep naaktzadigen (Gymnospermae) omvat een aantal ordes, waarvan veel alleen fossiel bekend zijn, zoals zaadvarens, de Cycadeoidales, de Pentoxylales en de Cordaitales. De groepen met recente vertegenwoordigers zijn de palmvarens, de Ginkgoales, de Coniferales en de Gnetales. Bij deze ordes is er, naast morfologisch onderzoek, ook cytologisch en biochemisch onderzoek mogelijk ten behoeve van het opstellen van een, weliswaar incomplete, fylogenetische stamboom. De slechts fossiel bekende groepen kunnen alleen worden ingedeeld en tussengevoegd op grond van hun bewaard gebleven morfologische kenmerken, in vergelijkend onderzoek met de recente groepen, waarvan de fylogenetische samenhang niet geheel duidelijk is.
Palmvarens, Cycadales[bewerken | brontekst bewerken]

De Cycadales of palmvarens, een orde in de klasse Cycadopsida[7], zijn betrekkelijk kleine struiken, bomen of klimplanten met geveerde bladen (macrofyllen).
De kiemplanten hebben twee zaadlobben. De volwassen planten zijn strikt tweehuizig, en hebben een soms knolvormige stam. De macrofyllen of bladen zijn enkel of dubbel veerdelig, waardoor veel planten een palmachtig uiterlijk hebben.
De macrosporangia staan op schubachtige macrosporofyllen in eindstandige of axillaire strobili (kegels). Bij Cycas staan de vrouwelijke sporofyllen niet in een strobilus, maar evenals de macrofyllen aan de top van de stam. Bij sommige Cycas-soorten zijn deze bladachtige macrosporofyllen duidelijke veernervig, bij andere soorten zijn ze sterk gereduceerd tot schubben en is de bladvorm niet meer herkenbaar.
De zaadknoppen zijn groot en met het blote oog goed zichtbaar en staan per 2 (tot 12 bij Cycas) op een macrosporofyl. De zaadknop heeft 1 dik integument, bestaande uit een beschermende, drielagige "testa". De macrospore in de zaadknop ontwikkelt zich tot macroprothallium. Boven het macroprothallium is er in de prothalliumkamer ruimte is voor de antherozoïden (zaadcellen). Het macroprothallium heeft twee tot enkele archegonia met een eicel. Op de micropyle, aan het uiteinde heeft de zaadknop, wordt een pollinatiedruppel gevormd, waarin het pollen blijft hangen.
De microsporofyllen dragen enkele tientallen tot honderden, in groepen (sori) bijeenstaande microsporangia. De door de wind verspreide microsporen worden opgevangen door een pollinatiedruppel. Ten tijde van de bestuiving ontwikkelt de microspore binnen de sporewand tot 3-cellige gametofyt: een prothalliumcel, een generatieve cel en een pollenbuiscel (de grootste cel). Men spreekt dan niet meer van microspore maar van pollenkorrel.
Bij het indrogen van de pollinatiedruppel wordt het pollen naar binnen gezogen naar de pollenkamer, waar de pollenkorrels kiemen door de vorming van een pollenbuis, die bij de nucellus naar binnen groeit. De generatieve cel deelt zich in een pollenbuiscel (steelcel) en een spermatogene cel. De mannelijke gametofyt bestaat uit een pollenbuis en heeft geen antheridia, maar vormt uit de spermatogene cel twee ongeveer ¼ mm grote spermatozoïden met in spiralen staande banden van korte flagellen hebben.
-
 Vrouwelijke sporofyl van Dioon edule met 2 zaden.
Vrouwelijke sporofyl van Dioon edule met 2 zaden. -
 Vrouwelijke sporofyllen van Cycas revoluta. Deze staan langs de stam.
Vrouwelijke sporofyllen van Cycas revoluta. Deze staan langs de stam. -
 Microsporofyllen van Encephalartos villosus. De microsporangia (= pollenzakken) zijn gedeeltelijk geopend.
Microsporofyllen van Encephalartos villosus. De microsporangia (= pollenzakken) zijn gedeeltelijk geopend.



Ginkgoales[bewerken | brontekst bewerken]
De Ginkgoales zijn een orde in de klasse Coniferopsida.[8] Er is slecht 1 recente soort, Ginkgo biloba (Japanse notenboom) in de familie Ginkgoaceae, die mogelijk wild voorkomt in China. Het zijn strikt tweehuizige bomen, met lange en korte loten. De bladen hebben dichotoom vertakte bladnerven. De bladeren vertonen bladdimorfie: aan de lange loten zijn de bladen wat dieper ingesneden dan die aan de korte loten. Het embryo heeft twee kiemlobben en geen duidelijke suspensor.
De microsporangia staan in strobili met zijasjes (ook wel sporangioforen) die elk 2 microsporangia dragen. De ontwikkeling van de microspore bij Ginkgo biloba vertoont een sterke overeenkomst met die bij de den (Pinus).[9]
De ovula (zaadknoppen) staan in paren op een dichotoom vertakte as, met de eindstandige macrosporangia. Binnen de integumenten heeft de nucellus aan de top een pollenkamer. Door meiose ontstaat er een lineaire tetrade van macrosporen, waarvan alleen de eerste zich ontwikkelt tot een chlorofyl bevattend macroprothallium (gametofyt). Hier groeien twee tot drie archegonia met een eicel.
- Japanse notenboom
-
 Mannelijke kegels
Mannelijke kegels -
 Vrouwelijke bloeiwijzen
Vrouwelijke bloeiwijzen -
 Zaden
Zaden



Coniferen, Coniferales[bewerken | brontekst bewerken]
De Coniferales zijn gewoonlijk forse bomen met pycnoxyl secundair hout. De naaldvormige bladen zijn primair macrofyllen met dichtoom vertakte nerven, maar secundair 1-nervige microfyllen.[10]
De voortplantingsorganen van de Coniferales (coniferen) zijn divers van bouw. De kegels met de sporangia zijn eenslachtig.
Bij Pinus (den) heeft de mannelijke strobilus schubvormig microsporofyllen met 2-15 pollenzakjes aan een schub.
De microsporemoedercellen vormen de microsporen, die blaasvormige uitstulpingen hebben. Het microprothallium bestaat uit microprothalliumcellen en een pollenbuiscel, waarnaast nog 2 generatieve kernen worden gevormd. Vorming van de mannelijke gameten bij de Coniferales bij de families Pinaceae, Podocarpaceae, Araucariaceae; bij Cephalotaxaceae, Taxodiaceae en Cupressaceae gaat de ontwikkeling na de vorming van de kern van de microspore verder met de vorming van de pollenbuiskern en de generative cel.[11]
De vrouwelijke kegels zijn samengesteld: er is een hoofdas met vele tot enkele schubvormige schutbladen. De zaadknoppen-dragende schub kan daarmee in mindere of meerdere mate vergroeid zijn. De bevruchting vindt plaats door windbestuiving en sifonogamie: de pollenbuis groeit naar de eicel, waar de spermakern versmelt met de eicel. Er vinden meer bevruchtingen plaats, zodat er meer embryo's ontstaan (dit verschijnsel heet polyembryonie). Slechts één embryo per zaadknop ontwikkelt zich uiteindelijk. De kiemplanten hebben tot 15 zaadlobben (cotylen).
- Verscheidenheid bij Coniferales
-
 Juniperus communis (Jeneverbes),
Juniperus communis (Jeneverbes),
tak met vruchten -
 Taxus baccata (Venijnboom),
Taxus baccata (Venijnboom),
detail takje met vruchten -
 Podocarpus polystachys,
Podocarpus polystachys,
bladeren -
 Cephalotaxus harringtonia,
Cephalotaxus harringtonia,
takken met vruchten -
 Pinus sylvestris (Grove den),
Pinus sylvestris (Grove den),
lang lot met 2 zaadkegels tussen bebladerde korte loten -
 Larix decidua (Europese lork),
Larix decidua (Europese lork),
takje met bebladerde korte loten -
 Taxodium distichum (Moerascipres),
Taxodium distichum (Moerascipres),
in natuurlijke omgeving -
 Podocarpus macrophyllus met vruchten
Podocarpus macrophyllus met vruchten
.JPG)







Gnetales[bewerken | brontekst bewerken]

groen – Welwitschia
blauw – Gnetum
rood – Ephedra
purper – Gnetum en Ephedra overlap verspreidingsgebied
Tot de orde Gnetales behoren drie geslachten, elk in een eigen familie: de Gnetaceae met Gnetum, de Ephedraceae met Ephedra en de Welwitschiaceae met Welwitschia.[12]
Belangrijke kenmerken van de orde zijn de houtvaten (tracheeën) in het secundaire hout, waarschijnlijk door parallelle evolutie uit tracheïden.
De planten zijn gewoonlijk tweehuizig. De voortplantingsorganen staan gegroepeerd in samengestelde strobili, die hier ook wel "bloeiwijzen" genoemd worden. De "bloemen" zijn meestal eenslachtig.
Vaak zijn er om het ovulum verschillende omhulsels, die geïnterpreteerd kunnen worden als periant of extra integumenten. De vrouwelijke "bloemen" hebben een enkele, rechtopstaande zaadknop, waarvan de nucellus is omgeven door twee of drie omhulsels: twee integumenten en een "periant". Een micropyle is de opening van het integument dat de nucellus (macrosporangium) omgeeft.
De mannelijke "bloem" bestaat uit een "periant" (bloemdek) met daarbinnen antheroforen met elk 2 tot 8 synangia (ook wel "pollenzakjes") - een tot een samenhangend geheel vergroeide sporangia.
De micropyle groeit bij de Gnetales uit tot een lang uittredende micropylaire buis, waardoorheen een druppel vocht naar buiten wordt gevormd om de stuifmeelkorrels in de lucht op te kunnen vangen. Bij indrogen van het vocht worden de stuifmeelkorrels naar binnen gezogen. De wijze van bestuiving wordt het "druppelmechanisme" genoemd. De bevruchting vindt plaats door de vorming van een pollenbuis met twee mannelijke gameetkernen.
In de nucellus vormen zich door meiotische deling uit een sporemoedercel de vier haploïde macrosporen in een tetrade. Hiervan gaan er drie te gronde. De overgebleven macrospore (of ten minste de kern van de macrospore) deelt zich meermaals tot enkele of meerdere cellen of celkernen: het macroprothallium of macrogametofyt. Ook dit wordt wel "embryozak" genoemd. Een van de celkernen van de embryozak functioneert als eicel.
Het embryo heeft twee kiembladen (cotylen).
Fylogenie[bewerken | brontekst bewerken]
Fylogenetische stamboom van de Archaeplastida, vereenvoudigd schema
|
|---|
|
|
- ↑ (en) Rubinstein, C.V., Gerrienne, P., De La Puente, G.S. (2010). Early Middle Ordovician evidence for land plants in Argentina (eastern Gondwana). New Phytologist 188 (2): 365–9. PMID 20731783. DOI: 10.1111/j.1469-8137.2010.03433.x.
- ↑ Tolweb plaatst Spermatopsida in de "Embryophytes" geconsulteerd 22 oktober 2006.
- ↑ Siebel, H.N. & H.J. During (2006). Beknopte mosflora van Nederland en België. KNNV Uitgeverij
- ↑ In Engelstalige botanische literatuur wordt, mogelijk in verband met de uitspraak, in plaats van het voorvoegsel macro- gewoonlijk het voorvoegsel mega- gebruikt. Voorbeelden zijn: megaspore, megaprothallium. In de overige Europese botanische literatuur is dit echter ongebruikelijk.
- ↑ Verklaring kleuren en randen:
- dikkeranden: meercellige generatie
- dunneranden: eencellig stadium
- ↑ De kleuren groen en oranjerood worden in de schema's gebruikt voor de ♀ gameten, resp. voor de ♂ gameten.
- ↑ Kalkman, C. (1972) p 142, 160, 195
- ↑ Kalkman C. (1972) p. 169 e.v.
- ↑ Sporne, K.R. (1965) p. 170.
- ↑ Kalkman, C. (1972) p. 195
- ↑ Sporne, K.R. (1965) p. 142.
- ↑ EMBRYOPSIDA Pirani & Prado
- ↑ ANA grade: Amborellales, Nymphaeales en Austrobaileyales
Literatuur
- DESCRIPTION OF THE SUPERGROUP UNIKONTA (Cavalier-Smith et al. 2002) Geraadpleegd: 15-05-2017.