
Ascomyceten
| Ascomyceten Fossiel voorkomen: Vroeg-Devoon[1] – heden | |||
|---|---|---|---|

| |||
| Houtknotszwam (Xylaria polymorpha) | |||
| Taxonomische indeling | |||
| |||
| Stam | |||
| Ascomycota Cavalier-Smith (1998) | |||

| |||
| Groot takmos (Ramalina fraxinea) met apotheciën | |||
| Afbeeldingen op | |||
| Ascomyceten op | |||
| |||
Ascomyceten (Ascomycota), ook wel zakjeszwammen genoemd, zijn een stam in het rijk der schimmels. De ascomyceten vormen een grote groep met meer dan 64.000 beschreven soorten.[2][3] Ze danken de naam zakjeszwammen aan de sporenzakjes ('ascus', meervoud 'asci'), de karakteristieke voortplantingsstructuren die microscopisch waarneembaar zijn. Veel bekende schimmels, waaronder de gisten en de korstmossen, worden tot deze groep gerekend, evenals enkele eetbare paddenstoelen, zoals morieljes en truffels.
Zakjeszwammen hebben zich naar alle waarschijnlijkheid ontwikkeld als zustergroep van de steeltjeszwammen (Basidiomycota) en vormen nu samen daarmee het onderrijk Dikarya.[4] Een belangrijk verschil tussen de twee groepen is dat bij een zakjeszwam sporen inwendig gevormd worden in een sporenzakje en bij een steeltjeszwam sporen uitwendig gevormd worden aan een specifieke cel (basidium).
Vroeger werden schimmels waarvan alleen het ongeslachtelijke stadium (het anamorfe stadium) bekend is in een restgroep ingedeeld, de Deuteromycota. Inmiddels kan de meerderheid van deze schimmels op grond van moleculairgenetische gegevens tot de zakjeszwammen gerekend worden.
Zakjeszwammen spelen een belangrijke rol in veel ecosystemen, bijvoorbeeld als mycobiont (schimmelpartner) in korstmossen, endofyten en mycorrhizas, en bovendien als reducent. De vruchtlichamen van zakjeszwammen vormen voedsel voor verschillende dieren, van insecten en slakken tot knaagdieren en grote zoogdieren als reeën en wilde zwijnen. Ook zijn er zakjeszwammen die parasiteren op andere schimmels. Zo parasiteert Eudarluca caricis onder andere op Pucciniaceae-soorten.[5]
Zakjeszwammen zijn ook voor de mens van grote betekenis. In negatieve zin doordat ze verantwoordelijk zijn voor talrijke ziektes bij mensen, huisdieren en cultuurgewassen. In positieve zin omdat ze een belangrijke rol spelen bij de bereiding van levensmiddelen als kaas, brood, bier en wijn, en omdat ze van grote waarde zijn voor de productie van medicijnen. Zo heeft het antibioticum penicilline, dat geproduceerd wordt uit Penicillium chrysogenum, in de tweede helft van de twintigste eeuw geleid tot een omwenteling in de bestrijding van bacteriële infectieziekten.
Bouw[bewerken | brontekst bewerken]


Veel zakjeszwammen hebben de vorm van een wollige zwamvlok of mycelium. Het mycelium bestaat uit lange, zich vertakkende schimmeldraden of hyfen, die ongeveer vijf micrometer dik zijn. Een mycelium wordt meestal enkele centimeters groot, maar de schimmeldraden daarin zijn kilometers lang. Het tegengestelde van deze vormen zijn de eencellige gisten die vaak alleen met een microscoop waargenomen kunnen worden. Vermeldenswaard is ook dat er ascomyceten zijn die als mycelium en als eencellige voorkomen. Voorbeelden zijn Histoplasma capsulatum en Blastomyces dermatitidis. Een aantal gisten zoals bakkersgist kan zowel in een eencellige als meercellige vorm voorkomen.
Bij de meeste zakjeszwammen bestaat de celwand uit chitine en β-glucaan. De schimmeldraden zijn op karakteristieke wijze in compartimenten verdeeld door septa (interne wanden). Deze geven de schimmeldraad stevigheid en verhinderen grootschalig verlies van cytoplasma als in de celwand een breuk optreedt. De meeste septen hebben in het midden een kleine opening waardoor celplasma en ook celkernen zich min of meer vrij door de hele schimmeldraad kunnen bewegen. Bij de opening liggen een tot vijf kleine, ronde, lichtbrekende lichaampjes, de zogenoemde Woronin-lichaampjes, die de opening bij breuk afsluiten, zodat de celinhoud van de aanliggende compartimenten niet wegstroomt. Schimmeldraden hebben doorgaans slechts één kern per cel, men noemt ze dan 'uninucleaat'.
Veel zakjeszwammen hebben een met het blote oog zichtbaar "vruchtlichaam", het ascocarp, dat een functie heeft bij de geslachtelijke voortplanting. De term vruchtlichaam is weliswaar zeer gebruikelijk, maar heeft niets te maken met een vrucht zoals die voorkomt bij planten. Een ascocarp ligt, meestal met meerdere tegelijk, in een stroma, een dichte structuur van vegetatieve, vaak verdikte en sterk gepigmenteerde schimmeldraden, die een plectenchymatisch tot pseudoparenchymatisch weefsel vormen. Soms is het stroma overtrokken met sporen (conidiën).[6] Het op een paddenstoel lijkend stroma met daarin de ascocarpen komt onder andere voor bij de eetbare truffel, de morielje en de daarop lijkende, giftige voorjaarskluifzwam.
Sommige zakjeszwammen, behorend tot de Taphrinales, vormen geen ascocarp, zoals Taphrina deformans dat de perzikkrulziekte veroorzaakt. Alle cellen van het mycelium van deze zakjeszwammen hebben twee kernen. De sporenzakjes worden gevormd door een proascus en zitten op het blad.
De Ascomyceten als groep hebben als gemeenschappelijk kenmerk het sporenzakje (ascus).[7] De sporenzakjes hebben niet alleen een van de andere cellen afwijkende bouw, maar hebben ook een eigen functie. In het ascus vindt de karyogamie (kernversmelting) plaats als laatste stap van de bevruchting, maar direct hierop volgt nog de meiose waarbij de vier ascosporen worden gevormd. Vaak volgt nog een gewone celdeling (mitose), waardoor er acht cellen aanwezig zijn in het sporenzakje. Verder zijn er bij sommige typen sporenzakjes verschillende mechanismen te vinden in de asci voor het actief verspreiden van de sporen.
Ascocarp[bewerken | brontekst bewerken]

Het ascocarp bestaat uit parafysen en fertiele hyfen. Parafysen zijn lange, steriele organen, die tussen de sporenzakjes staan. De parafysen vormen samen het hamathecium. De hyfen in het ascocarp zijn zeer sterk ineengevlochten en vormen de voortplantingscellen. Ze ontwikkelen zich meestal op het binnenste oppervlak van de ascocarp en staan doorgaans in een eigen vruchtlaag, het hymenium, waarin de cellen zich ontwikkelen tot sporenzakjes.
Men deelt de ascocarpen onder andere in naar hun ligging: bevindt het ascocarp zich bovengronds zoals bij de morielje, dan noemt men ze 'epigeïsch'. Ligt het ondergronds zoals bij truffels, dan heten ze 'hypogeïsch'. Deze typologie heeft overigens geen grote betekenis voor de indeling van de zakjeszwammen.
Het ascocarp kent meerdere verschijningsvormen die als volgt beschreven worden:
Apothecium[bewerken | brontekst bewerken]
Het apothecium is een bekerachtig, naar boven geopend ascocarp. De vruchtlaag (hymenium) ligt vrij, zodat gelijktijdig talrijke sporen verspreid kunnen worden. Voorbeelden van zakjeszwammen met apotheciën zijn morieljes en kluifzwammen.
-
 Voorbeeld van een apothecium, met daarin aangegeven de meiotische ontwikkeling van asci (paars), rijpe asci (blauw) en parafysen (rood)
Voorbeeld van een apothecium, met daarin aangegeven de meiotische ontwikkeling van asci (paars), rijpe asci (blauw) en parafysen (rood) -
 Het korstmos groot dooiermos met apotheciën.
Het korstmos groot dooiermos met apotheciën. -


.jpg)

Cleistothecium[bewerken | brontekst bewerken]
Het cleistothecium (chasmothecium) is een gesloten bolvormig ascocarp. De sporen kunnen daardoor niet zelfstandig vrijkomen. Schimmels met cleistotheciën hebben verschillende strategieën voor verspreiding van hun sporen. Truffels bijvoorbeeld lokken dieren aan, zoals wilde zwijnen, die het ascocarp openbreken en de daaraan aangehechte sporen over grote afstanden kunnen verspreiden. Ook regendruppels kunnen de celistotheciën openbreken.
Cleistotheciën worden onder andere aangetroffen bij schimmels die slechts zeer weinig ruimte voor hun ascocarp tot hun beschikking hebben, zoals bij soorten die onder de schors van bomen leven, of in de bodem liggen. Echte meeldauw-soorten hebben de cleistotheciën op het oppervlak van het blad zitten. Ook de huidschimmels van het geslacht Trichophyton, waartoe onder andere Trichophyton rubrum behoort, vormen cleistotheciën.
Het onrijpe cleistothecium zit met hyfen verankerd in het mycelium. Deze hyfen vormen later de aanhangsels van het cleistothecium.
-
 Opengebroken cleistothecium van Pseudallescheria boydii
Opengebroken cleistothecium van Pseudallescheria boydii -
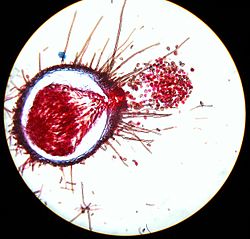 Zich openend cleistothecium van echte meeldauw
Zich openend cleistothecium van echte meeldauw -
 Cleistothecium met ascosporen van echte meeldauw
Cleistothecium met ascosporen van echte meeldauw -
 Cleistothecium van eikenmeeldauw op een blad van de zomereik. De aanhangsels van het cleistothecium zijn per schimmelsoort verschillend.
Cleistothecium van eikenmeeldauw op een blad van de zomereik. De aanhangsels van het cleistothecium zijn per schimmelsoort verschillend.




Perithecium[bewerken | brontekst bewerken]
Een perithecium is een hard, min of meer bolrond of flesvormig, gesloten ascocarp. Kenmerkend is dat zich aan de bovenzijde een kleine porie, de 'ostiole' bevindt, waardoorheen de sporen bij rijpheid een voor een vrijgelaten worden, dit in tegenstelling tot hetgeen bij apotheciën het geval is. Peritheciën komen onder andere voor bij Xylaria zoals de geweizwam, bij Nectria-soorten zoals het gewoon meniezwammetje en bij Claviceps-soorten, zoals moederkoorn.
-
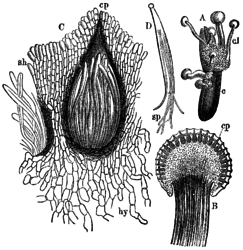 A: Gekiemd sclerotium van moederkoorn met ascocarp, B:Stroma met peritheciën, C:Perithecium met asci, D:Ascus met draadvormige sporen.
A: Gekiemd sclerotium van moederkoorn met ascocarp, B:Stroma met peritheciën, C:Perithecium met asci, D:Ascus met draadvormige sporen. -
 Stroma met peritheciën van moederkoorn
Stroma met peritheciën van moederkoorn -
 Peritheciën van het gewoon meniezwammetje
Peritheciën van het gewoon meniezwammetje -
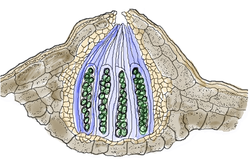 Doorsnede van een perithecium
Doorsnede van een perithecium





Pseudothecium[bewerken | brontekst bewerken]
Een pseudothecium is een variant van het perithecium. Het onderscheid bestaat uit de manier waarop de sporen vrijkomen: in pseudotheciën worden dubbelwandige sporenzakjes gevormd. Als de zakjes nat worden nemen ze water op en zetten uit, waardoor de sporen uit de opening worden geschoten. Voorbeelden zijn appelschurft en Didymella bryoniae.
Gymnothecium[bewerken | brontekst bewerken]

Het gymnothecium is een naakte ascocarp, zo genoemd omdat het niet bedekt wordt door een beschermende laag, zoals een peridium of een pseudoparenchymatisch weefsel. Bij Myxotrichum vormen de buitenste hyfen borstels, de zogenaamde setae. Deze bestaan uit stralen dikwandige, gekleurde, stijve hyfen. Als het sporenzakje bewogen wordt door bijvoorbeeld wind, water, mijten of insecten, vallen de sporen door openingen tussen de stralen van het hyfenvlechtwerk naar buiten.[* 1][* 2][* 3] Een ander voorbeeld van een zakjeszwam die een gymnothecium vormt is Blastomyces dermatitidis, die blastomycose bij mensen veroorzaakt.
Sporocyst[bewerken | brontekst bewerken]
De sporocyst is een ascocarp dat voorkomt bij zakjeszwammen van de familie Ascosphaeraceae in de klasse Eurotiomycetes.[8] De sporocyst wordt gevormd uit een enkele cel, de nutriocyst, is eencellig en cyste-achtig en heeft een dubbele wand. De sporenzakjes liggen in groepen in sporenbollen, die elk omgeven zijn door één membraan. Een sporenbol kan twee tot honderden sporenzakjes bevatten. De sporenzakjes komen vrij in de sporocyst te liggen doordat de wand van de sporenbol in stukjes breekt.
-
 Ascosphaera fimicola: A: twee iriserende sporocysten nog vast aan de hyfen. B–C: close-up van sporocysten. D: ascosporen. E: sporenbol.
Ascosphaera fimicola: A: twee iriserende sporocysten nog vast aan de hyfen. B–C: close-up van sporocysten. D: ascosporen. E: sporenbol. -
 Ascosphaera callicarpa: B: larve bedekt met hyfen en sporocysten. C en D: sporocysten. E: ascosporen.
Ascosphaera callicarpa: B: larve bedekt met hyfen en sporocysten. C en D: sporocysten. E: ascosporen.


Sclerotium[bewerken | brontekst bewerken]

De zwamvlok (mycelium) kent een rustfase waarin hij een vorm aanneemt waarin hij kan overleven: het sclerotium. Een sclerotium is rijk aan vet en koolhydraten en bestaat uit een aan elkaar vastgeklitte schimmeldraden (pseudoparenchymatisch weefsel). In de volksmond noemt men dat een 'wolventand' of 'rattenkeutel'.
De sclerotiën vallen op de grond en kunnen na vijf tot tien jaar nog kiemen, een apothecium vormen en vervolgens planten aantasten. Of dit gebeurt is sterk afhankelijk van het weer. Het sclerotium moet zich in een vochtige omgeving bevinden en de temperatuur moet bij voorkeur tussen de 10 en 20 °C liggen. In het voorjaar kunnen de sclerotiën al gaan kiemen als de temperatuur boven de 5 °C komt en er gedurende tien dagen vochtige omstandigheden zijn.
Ecologische rol[bewerken | brontekst bewerken]
Zakjeszwammen spelen een centrale rol in de meeste ecosystemen op het land. Ze zijn belangrijke reducenten die organische materialen omzetten in anorganische stoffen. Omdat ze ook moeilijk afbreekbare substanties zoals cellulose, lignine en keratine kunnen reduceren, zijn ze van groot belang voor de stikstof- en koolstofkringloop.
Daarnaast hebben zakjeszwammen talrijke symbiotische relaties met andere organismen. Het bekendst zijn relaties met groenwieren en blauwwieren (Cyanobacteria) in korstmossen. Twee andere vormen van symbiose met planten zijn relaties met wortels (mycorrhiza) en met andere delen van planten (endofyt).
Als saprobiont[bewerken | brontekst bewerken]
Veel soorten zakjeszwammen leven als saprobiont: ze voeden zich met organisch afval als dorre bladeren, dode takken, afgestorven bomen en dode dieren. Ook collageen (een dierlijk structuureiwit) of keratine (waaruit haar en nagels bestaan) kunnen als voedingsbron dienen. Saprobionte zakjeszwammen produceren verteringsenzymen die de organische verbindingen afbreken. De afbraakproducten worden vervolgens door de celwand van de zwam opgenomen. Een dergelijke levenswijze noemt men chemotroof en chemotrofe soorten worden gerekend tot de reducenten.
Als parasiet[bewerken | brontekst bewerken]
Andere zakjeszwammen parasiteren op planten, dieren of andere schimmels en dragen zo bij aan het ecologisch evenwicht. Ze kunnen er als sleutelsoort voor zorgen dat een bepaalde soort niet de overhand krijgt, zodat verschillende soorten naast elkaar kunnen blijven bestaan. Een dergelijke levenswijze noemt men heterotroof. Een groep zakjeszwammen, de entomopathogene schimmels, tasten insecten aan, waaronder bladsnijdersmieren. Voorbeelden daarvan zijn soorten die tot de geslachten Cordyceps, Metarhizium en Ophiocordyceps behoren. Bijzonder aan deze groep zijn enkele extreme specialisaties; zo tasten sommige soorten zakjeszwammen slechts één bepaalde poot van één bepaalde insectensoort aan.
|







_A._Massal_243169.jpg)

Als symbiont[bewerken | brontekst bewerken]
Korstmossen[bewerken | brontekst bewerken]
Korstmossen zijn zakjeszwammen die in mutualistische symbiose leven met groenwieren of blauwalgen, soms ook met beide. De symbionten worden respectievelijk 'mycobiont' (de schimmel) en 'fycobiont' (het groenwier en/of de blauwalg) genoemd. Ongeveer 18 000 van alle zakjeszwammen zijn korstmosvormend; dat is 42 procent van het totaal. Omgekeerd zijn de schimmelpartners van vrijwel alle korstmossen zakjeszwammen - het aandeel van steeltjeszwammen ligt waarschijnlijk op niet meer dan twee tot drie procent. Zakjeszwammen hebben waarschijnlijk reeds vroeg in hun ontstaansgeschiedenis groenwieren, blauwalgen en andere algen "gedomesticeerd" en vormen met deze de als korstmossen bekendstaande leefgemeenschappen.
Korstmossen kunnen leven op de meest extreme biotopen, zoals in poolgebied, woestijn en hooggebergte. Ze kunnen overleven bij temperaturen van -40 tot +80 graden Celsius. De samenwerking bestaat eruit dat de wieren of algen door fotosynthese de stofwisselingsenergie aan de schimmels beschikbaar stellen, en de schimmels een stabiel ondersteuningsskelet bieden en beschermen tegen straling en uitdroging.
Korstmossen kunnen gecompliceerde secundaire metabolieten produceren, die de symbionten niet zelfstandig kunnen maken. Een voorbeeld is usninezuur, een dibenzofuraan dat in de natuur voorkomt bij epifytische korstmossen. Usninezuur wordt aangetroffen in veel verschillende geslachten, zoals Usnea (baardmos), Cladina (rendiermos), Lecanora, Ramalina, Evernia, Parmelia en Alectoria.
Mycorrhiza's[bewerken | brontekst bewerken]
Mycorrhiza's zijn zakjeszwammen die symbiotisch verbonden zijn met wortelstelsels van planten. Mycorrhiza's zijn voor veel planten van levensbelang omdat ze hen toegang bieden tot voedingsstoffen in de bodem. De schimmel, die door zijn fijn vertakt hyfenweefsel veel beter in staat is mineralen uit de bodem op te nemen dan de plant zelf, wordt op zijn beurt door de plant voorzien van fotosyntheseproducten. Het is bekend dat wortelschimmels zelfs in staat zijn door hun wijdvertakte hyfenweefsel voedingsstoffen van de ene plant naar de andere te transporteren. Waarschijnlijk maakten relaties tussen wortelschimmels en plantenwortels uiteindelijk de verovering van het land vanuit de zee mogelijk. In ieder geval zijn de alleroudste fossielen van landplanten met wortelschimmels geassocieerd, wat wil zeggen dat de schimmels als endosymbiont gevonden zijn in fossielen.[9]
Endofyten[bewerken | brontekst bewerken]
Endofyten zijn zakjeszwammen die leven in een waardplant, niet in de wortels, maar in het bijzonder in de stengel en in de bladeren. Men onderscheidt endofyten die alleen plaatselijk in de plant voorkomen en endofyten die in de gehele plant leven. Endofyten beschadigen de waardplanten gewoonlijk niet. De relatie tussen schimmel en waardplant is nog niet goed begrepen: het schijnt dat een plant die endofyten heeft een hogere weerstand tegen schadelijke insecten, rondwormen en bacteriën krijgt. Besmetting van een plant met endofyten leidt tot de productie van speciale alkaloïden, die bij plantenetende zoogdieren de gezondheid schaden.
Symbiose met dieren[bewerken | brontekst bewerken]


Een aantal soorten zakjeszwammen uit het geslacht Xylaria wordt aangetroffen in nesten van bladsnijdersmieren en verwante insectensoorten uit de geslachtengroep Attini, en in de schimmeltuinen van termieten.
Belangrijke symbionten zijn verder de schorskevers. De keverwijfjes transporteren de schimmelsporen in uitstulpingen van het voorste rugschild, het pronotum, naar nieuwe planten. Doorgaans vreten ze in het hout gangen die zich naar binnen tot grotere ruimten verbreden. In deze kamers leggen ze hun eieren die zich daar ontwikkelen tot larven. De schimmelsporen die zijn achtergelaten vormen hyfen, die het hout enzymatisch afbreken zodat het kan dienen tot voedsel voor de larven. Na hun verpopping en metamorfose verspreiden de volwassen insecten de schimmelsporen naar nieuwe planten. Een bekend voorbeeld is de iepenziekte (Ophiostoma ulmi), die door de iepenspintkevers (Scolytus scolytus en Scolytus multistriatus) wordt verspreid.
Als carnivoor[bewerken | brontekst bewerken]
Ten slotte zijn er vleesetende schimmels waaronder de eerder genoemde mycorrhiza's,[10] en nematofage schimmels, die met hun hyfen vallen vormen, waarin ze kleine protisten zoals amoeben, maar ook meercellige dieren als rondwormen,[11][12][13] raderdieren, beerdiertjes of zelfs geleedpotigen zoals springstaarten kunnen vangen. Ze behoren tot de familie Orbiliaceae.
Er zijn zes verschillende vormen van vallen: samentrekkende ringen, niet samentrekkende ringen, zittende kleverige knobbels, gesteelde kleverige knobbels, kleverige netten en kleverige kolommen.[14]
Bijzondere substraten[bewerken | brontekst bewerken]
Een voorbeeld van bijzondere substraten die door zakjeszwammen kunnen worden gereduceerd is kerosine, dat door de kerosineschimmel (Amorphotheca resinae) wordt afgebroken. Deze schimmel komt voor in brandstoftanks van vliegtuigen en schepen en kan leiden tot verstopte brandstofleidingen.[15]
Verspreiding en biotoop[bewerken | brontekst bewerken]
Zakjeszwammen als stam komen wereldwijd voor, in alle ecosystemen op het land. Een kleine groep is naar het water teruggekeerd. De verspreiding over land varieert sterk per soort. Sommige komen wereldwijd voor, als korstmossen zelfs in poolgebieden, woestijnen en hooggebergtes. Andere soorten hebben slechts een regionale verspreiding, zoals de culinair zeer begeerde witte truffel die alleen gevonden wordt in geïsoleerde streken in Italië en Frankrijk. Volgens het Nederlands Soortenregister komen van de 64 000 bekende soorten zakjeszwammen rond 3 000 soorten in Nederland voor.[16]
Doordat bij de meeste zakjeszwammen de celwand uit chitine en β-glucaan bestaat en de schimmeldraden afsluitbare tussenwanden (septa) hebben, zijn ze in staat zich in drogere biotopen te vestigen dan bijvoorbeeld de lagere schimmels. Deze uitdrogingsresistentie is bij bepaalde soorten die op gezouten vis groeien, wegens het sterk hypertone milieu, extreem zichtbaar.
Voortplanting[bewerken | brontekst bewerken]
De voortplanting van zakjeszwammen kan zowel ongeslachtelijk als geslachtelijk plaatsvinden. De laatste manier is bijzonder karakteristiek door de zichtbare voortplantingsstructuren, de sporenzakjes. Aan de andere kant speelt de ongeslachtelijke voortplanting veruit de grootste rol; veel soorten hebben de geslachtelijke voortplanting geheel opgegeven (of deze werd nog niet waargenomen).
Men onderscheidt bij zakjeszwammen twee fundamenteel verschillende fasen in de levenscyclus, het 'anamorf' of ongeslachtelijke stadium, en het 'teleomorf' of geslachtelijke stadium, wat dus niet altijd meer voorhanden is. Anamorf en teleomorf worden veelal gezamenlijk als 'holomorf' aangeduid.
Omdat het uiterlijk van de zakjeszwam in het ongeslachtelijk stadium sterk verschilt van dat in de geslachtelijke fase was het tot ver in de 20e eeuw vaak niet mogelijk de koppeling te zien tussen beide stadia. Als gevolg daarvan werd één schimmel vaak in twee verschillende soorten ingedeeld, onder twee namen. Zo is de geslachtelijke verschijningsvorm van de kerosineschimmel onder de naam Amorphotheca resinae bekend, terwijl de schimmel in het ongeslachtelijke stadium Hormoconis resinae wordt genoemd. Deze ongewenste situatie wordt in 2016 nog steeds taxonomisch geaccepteerd. Met behulp van moleculairgenetische methoden wordt bij steeds meer van deze "soorten" de koppeling ontdekt, zodat vroegere indelingen van het ongeslachtelijke stadium in een eigen afdeling, de Fungi imperfecti (Deuteromycota), gaandeweg overbodig worden.

Ongeslachtelijke voortplanting[bewerken | brontekst bewerken]
Ongeslachtelijke voortplanting is bij zakjeszwammen de dominante vorm van vermeerdering. Voortplanting vindt plaats door 'conidiën', ook wel 'conidiosporen' genoemd, ter onderscheiding van geslachtelijk ontstane sporen, de 'ascosporen'. Een andere term die gebruikt wordt is 'mitosporen', naar hun ontstaan door het cellulaire proces mitose. Deze sporen zijn eenkernige structuren die genetisch identiek zijn aan de zich voortplantende schimmel. Ze worden in de regel aangemaakt door speciale als 'conidiogeen' aangeduide cellen, die zich bevinden aan de uiteinden van gespecialiseerde schimmeldraden, de 'conidioforen'. Afhankelijk van de soort worden de sporen verspreid door wind, door water of door dieren.
- Conidioforen met sporen
-
 Conidioforen van Trichoderma fertile met vaasvormige phialiden en nieuw gevormde conidiën aan de toppen (lichte puntjes).
Conidioforen van Trichoderma fertile met vaasvormige phialiden en nieuw gevormde conidiën aan de toppen (lichte puntjes). -
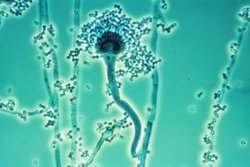 Conidioforen van soorten uit het geslacht Aspergillus, conidiogenese is blastisch-fialidisch
Conidioforen van soorten uit het geslacht Aspergillus, conidiogenese is blastisch-fialidisch -
 Conidioforen van Trichoderma harzianum, conidiogenese is blastisch-fialidisch
Conidioforen van Trichoderma harzianum, conidiogenese is blastisch-fialidisch -
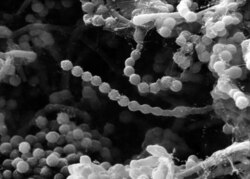 Conidioforen van Paecilomyces variotii (SEM-afbeelding)
Conidioforen van Paecilomyces variotii (SEM-afbeelding)




Coelomyceten en hyphomyceten[bewerken | brontekst bewerken]
De verscheidenheid van zakjeszwammen in het ongeslachtelijk stadium is enorm. De soorten laten zich (zonder betekenis voor de ordening) grofweg indelen in twee groepen, afhankelijk van de structuur waarin ze hun sporen maken (conidiomatum). Gebeurt dat in een afgesloten conidiomatum dan spreekt men van coelomyceten; is de structuur niet afgesloten heten ze hyphomyceten.

- Coelomyceten maken hun sporen aan in een afgesloten conidiomatum, die zich vormt vlak onder het oppervlak van een gastheerorganisme. Men onderscheidt:
- Pycnidiale conidiomata of 'pycnidia', die zich ontwikkelen in het weefsel van de schimmel. Ze hebben meestal de vorm van een buikige vaas; de sporen worden in de omgeving losgelaten door de ostiole, een opening aan het uiteinde van de vaas.
- Acervulare conidiomata of 'acervuli', die zich in tegenstelling tot pycnidia niet ontwikkelen in het schimmelweefsel, maar in de gastheer. Ze kunnen op verschillende plaatsen liggen: onder de cuticula (het buitenste waslaagje van een plant), in de epidermis (de buitenste cellenlaag, opperhuid), direct daaronder (subepidermaal) of zelfs onder talrijke lagen van gastheercellen. Meestal wordt er een vlakke laag met relatief korte conidioforen gevormd, waarin massaal sporen worden geproduceerd. De daardoor steeds sterker wordende druk leidt uiteindelijk tot het openbreken van epidermis en cuticula, waardoor de sporen vrijkomen.
- Bij de hyphomyceten liggen de conidioforen, die aan het uiteinde conidiënvormende cellen dragen, altijd vrij. Ze komen meestal geïsoleerd voor, soms echter in de vorm van een parallel gerichte bundel of kussenvormige massa. Hun conidiomata worden in het eerste geval als 'synnemataal', in het laatste geval als 'sporodochiaal' aangeduid.
Sporen[bewerken | brontekst bewerken]
Voor de verdere classificatie van de ongeslachtelijke fase van zakjeszwammen zijn de sporen van belang, die onderscheiden kunnen worden naar septering en vorm:

- 'amerosporen' zijn sporen zonder tussenwand (septum); deze komen het meest voor
- 'didymosporen' hebben één tussenwand
- 'fragmosporen' zijn sporen met twee of meer scheidingswanden die geordend zijn als de sporten van een ladder (transversaal)
- 'dictyosporen' hebben ook twee of meer tussenwanden die echter een netwerkachtige structuur hebben
- 'staurosporen' kennen stralenvormige armen vanuit een centraal lichaam
- 'helicosporen' hebben het uiterlijk van een spiraalvormig opgewonden veer
- 'scolecosporen' zijn zeer lang en wormvormig, met een lengte-breedteverhouding van meer dan 15:1
- 'chlamydosporen' zijn ongeslachtelijke rustsporen van gisten. Deze dikwandige sporen kunnen ongunstige omstandigheden, zoals droge of hete seizoenen, overleven. Chlamydosporen hebben een ronde vorm en een glad oppervlak. Ze zijn meercellig, waarbij de cellen via poriën in het septum met elkaar in verbinding staan.

- Chlamydosporen
-
 Chlamydosporen van Coccidioides immitis
Chlamydosporen van Coccidioides immitis -
 Chlamydospore van de gist Candida albicans
Chlamydospore van de gist Candida albicans


Conidiogenese[bewerken | brontekst bewerken]
Een verder belangrijk kenmerk van de zakjeszwam in de ongeslachtelijke fase is de manier waarop de sporen gevormd worden, de 'conidiogenese'. Deze komt overeen met de embryologie bij planten en dieren, en laat zich in twee fundamentele ontwikkelingsvormen indelen. De eerste is de 'blastische' conidiogenese, waarbij de sporen reeds waarneembaar zijn voordat ze afgescheiden worden. De tweede is de 'thallische' conidiogenese, waarbij zich eerst de tussenwand vormt en de zo afgescheiden cel zich tot een spore omvormt.
Deze beide grondvormen laten zich nog verder onderverdelen:
- Bij 'blastisch-acropetale' sporenvorming ontstaan de sporen door het steeds verder uitlopen van de uiteinden van de conidiogene schimmeldraad, zodat zich een lange ketting van sporen vormt, waarbij de jongste altijd aan het uiteinde ('apikaal') ligt. Vormen zich twee uitlopers aan een spore, dat dan ramoconidium genoemd wordt, dan vertakt de keten zich. Blastisch-acropetale sporenvorming is bijgevolg een variant van het normale groei van schimmeldraden.
- De 'blastisch-synchrone' conidiogenese kenmerkt zich door de talrijke sporen die zich gelijktijdig vormen op een centraal gelegen, meestal opgezwollen cel. Veelal ontstaan uit deze sporen acropetale ketens.
- De 'blastisch-sympodiale' ontwikkeling wordt gekarakteriseerd doordat zich na de sporenvorming achter de sporen, aan de uiteinden van de conidioforen, een nieuw uiteinde ontwikkelt, dat zijdelings ('sympodiaal' respectievelijk 'zymös') verder groeit. Dit uiteinde vormt op zichzelf uiteindelijk een spore, waaronder weer een nieuw uiteinde ontstaat. De conidiogene cellen worden bij dit proces onvermijdelijk langer en langer.
- Bijzonder opvallend is de 'blastisch-annellidische' vorm van sporenvorming: Nadat een spore zich gevormd en afgescheiden heeft, blijft er een ringvormig litteken achter aan het uiteinde van de conidiogene cel. Deze cel groeit door de ring heen en vormt dan het volgende spore, dat bij zijn afscheiding op zijn beurt weer een litteken achterlaat, zodat men aan het aantal ringen van een condiogene cel kan aflezen hoeveel sporen er gevormd zijn.
- Bij de 'blastisch-fialidische' vorming van sporen ontstaan voortdurend nieuwe sporen aan het open eind van de conidiogene cellen, die in dit geval 'fialiden' genoemd worden. Gezien de fialide zelf niet van vorm en lengte verandert, groeit de sporenketting net als bij de blastisch-acropetale ontwikkeling naar boven. In tegenstelling tot deze blastisch-acropetale ontwikkeling zijn hier echter de aan de 'condiogene locus' gevormde onderste sporen altijd de jongste. De ongeslachtelijke vorm van de Penicillium is een bekend voorbeeld van deze wijze van sporenvorming.
- Als variant van de blastisch-fialidische ontwikkelingsvorm kan de 'basauxische' vorming gelden. Hierbij bevinden de sporen van de keten die ontstaat zich in verschillende ontwikkelingsstadia: terwijl de sporen aan het bovenste eind van de schimmeldraad reeds uitgerijpt zijn, hebben de sporen onderaan zich nog nauwelijks van de conidiogene schimmeldraad gedifferentieerd.
- Geheel anders verloopt de 'blastisch retrogressieve' sporenvorming. Hierbij vormt zich aan het uiteinde van de conidiogene hyfe een spore die zich door een tussenwand van de schimmeldraad afsplitst. Daaropvolgend ontwikkelt zich direct daaronder een volgende spore, dat zich nu op zijn beurt door een septe van de rest van de hyfe afsplitst. Door meervoudige herhaling van dit proces vormen zich uit een enkele schimmeldraad van het uiteinde naar beneden voortdurend nieuwe sporen terwijl de hyfecel zichzelf voortdurend verkort.
- Anders dan bij de blastische vormen worden bij 'thallish arthrische' conidiogenese eerst de septen gevormd. Daartoe dient een hyfe volgroeid te zijn om dan op onregelmatige afstanden dubbele tussenwanden te vormen. De zo ontstane individuele cellen scheiden zich op een 'schizolytisch' genoemde wijze van elkaar af en beginnen zich te differentiëren, zodat een keten van korte cilindervormige sporen, de zogeheten 'arthroconidiën' ontstaat, die met elkaar verbonden lijken te zijn. Bij een variant van deze conidiogenese vormen zich alternerend cellen en sporen, de cellen degenereren, sterven af en maken zo de tussenliggende arthroconidiën door rhexolytische dehiscentie vrij.
- Ten slotte bestaat er nog de 'thallisch solitaire' ontwikkelingsvorm. Hier scheidt zich aan het eind van een conidiogene schimmeldraad een grote, buikige cel af, waarin zich dan talrijke transversaal geordende interne tussenwanden vormen. De cel differentieert zich nu tot een 'fragmospoor' dat in zijn geheel (rhexolytisch) afgescheiden wordt.

Dehiscentie[bewerken | brontekst bewerken]
Het laatste onderscheidende kenmerk is de 'dehiscentie', het proces waarin de sporen zich afscheiden. Dehiscentie kan op twee verschillende manieren geschieden. Bij de 'schizolytische' varianten vormt zich tussen de sporen een dubbele scheidingswand met centraal gelegen middellamellen. Als deze wand uiteenvalt worden de sporen losgelaten. Bij rhexolytische dehiscentie, zoals bij soorten van het geslacht Coccidioides, worden de sporen uitwendig verbonden door een celwand die op een gegeven moment degenereert, waardoor de conidiosporen vrij komen.
Heterokaryose en paraseksualiteit[bewerken | brontekst bewerken]
Bij veel zakjeszwammen bestaat in het geheel geen geslachtelijk stadium of is het bestaan ervan niet bekend. Er zijn twee manieren waarop in het ongeslachtelijk stadium genenmateriaal kan worden uitgewisseld, en zo de genetische verscheidenheid kan worden behouden: 'heterokaryose' en 'paraseksualiteit'. De eerste komt simpelweg tot stand door versmelting van twee schimmeldraden, elk van een ander individu. Als resultaat van dat proces, dat anastomose genoemd wordt, bevinden zich gewoonlijk meerdere celkernen in de zwamvlok, die van verschillende individuen stammen.
Paraseksualiteit[* 4] is een fenomeen waarbij twee haploïde cellen van verschillende schimmeldraden zonder eigenlijke seksuele handelingen met elkaar versmelten, waarna de celkernen fuseren zodat een verdubbeling van het aantal chromosomen ontstaat. Daarna vindt een complexe vorm van kerndeling plaats, waarbij het tot een overkruising, een uitwisseling van genetisch materiaal tussen twee overeenkomstige chromosomen komt.[* 5] Uiteindelijk ontstaan door deling weer twee haploïde celkernen, die zich beide genetisch van de twee uitgangskernen onderscheiden.
Geslachtelijke voortplanting[bewerken | brontekst bewerken]
De geslachtelijke voortplanting bij zakjeszwammen verschilt van die bij andere schimmels doordat de sporen worden gevormd in een voor zakjeszwammen kenmerkende structuur, het sporenzakje of ascus.[17] Het sporenzakje fungeert als een 'sporangium', waarin deze geslachtelijke ascosporen worden aangemaakt. Daarnaast zijn er de ongeslachtelijk gevormde conidiosporen, die voor de ongeslachtelijke voortplanting dienen.
Bij de geslachtelijke voortplanting komt een kortdurende diploïde fase voor, de zygote, met in de celkern twee exemplaren van elk chromosoom (2n). Deze fase wordt in de regel na korte tijd beëindigd door een meiose (reductiedeling of rijpingsdeling) in het sporenzakje. Bij de meiose worden in eerste instantie vier haploïde ascosporen gevormd, maar vaak volgt nog een deling, een mitose, zodat er zich acht ascosporen bevinden in het sporenzakje. Meercellige ascosporen kunnen zich vormen door verdere mitosen. Bij de zo uit een zygote gevormde ascosporen is het aantal chromosomen in de celkern gehalveerd, van het diploïde aantal (2n) naar het haploïde aantal (n).
Afgezien van uitzonderingen als soorten die behoren tot de Taphrinales en bakkersgist zijn alle zakjeszwammen in normale toestand haploïde. De celkernen bevatten van elk chromosoom slechts één exemplaar, wat ze gevoelig maakt voor mutaties.
Verloop van de seksuele sporenvorming[bewerken | brontekst bewerken]
De geslachtelijke voortplanting wordt ingeleid als twee haploïde schimmeldraden (hyfen) met bij elkaar passende paringstypen samenkomen. De betrokken schimmeldraden stammen uit hetzelfde type hyfenweefsel als de ongeslachtelijke sporen. De opeenvolgende stappen in het fusieproces van de twee hyfen zijn beschreven door Fincham en Day[18] en door Wagner en Mitchell.[19]
Of er versmelting plaatsvindt, hangt af van de herkomst van de schimmeldraden. Bij veel schimmels versmelten hyfen alleen als ze bij hetzelfde organisme horen. Er treedt dan zelfbevruchting op, een eigenschap die als 'homothallie' bekendstaat. Andere schimmels hebben een genetisch niet-identieke partner nodig en zijn daarmee 'heterothallisch'. Daarnaast moeten de twee betreffende hyfen ook nog passende paringstypen hebben. Deze zijn een bijzonderheid van schimmels en kunnen vergeleken worden met geslachten bij planten en dieren, met dat verschil dat er bij schimmels meer dan twee paringstypen kunnen zijn.

Bij verenigbaarheid vormen zich op de hyfen 'gametangiën', de bouwcellen voor de geslachtscellen (gameten), waarin zich talrijke celkernen verzamelen. Deze gametangiën specialiseren zich in twee typen. Uit de eerste type, 'ascogonium' genoemd, ontstaat de 'trichogyne', een zeer fijne schimmeldraad. Deze vormt een brug naar een gametangium van het tweede type, het 'antheridium'. Via deze brug kunnen celkernen uit het antheridium zich naar het ascogonium bewegen. Anders dan bij planten en dieren komt het na de celversmelting (plasmogamie) in de regel niet tot een onmiddellijke kernversmelting (karyogamie). In plaats daarvan ordenen de kernen die uit het antheridium zijn binnengekomen zich paarsgewijs met kernen van het ascogonium, waarbij ze zelfstandig naast elkaar elkaar blijven bestaan. Daarmee begint de 'dikaryofase' in de levenscyclus van de schimmel, waarin bij de celdeling de kernparen zich synchroon delen.
De dikaryofase lijkt een evolutionaire aanpassing, waardoor in die gevallen van een zeldzame ontmoeting tussen twee individuen, er sporen ontstaan van een zo groot mogelijke verscheidenheid. Door de herhaalde synchrone deling van de kernparen vermeerdert het genetische uitgangsmateriaal zich en ontstaan bij de recombinatie gedurende de meiose veel herschikkingen, waardoor een grote genetische diversiteit gerealiseerd wordt. Ook steeltjeszwammen kennen een dikaryofase, reden waarom ze wel met zakjeszwammen worden samengevoegd in het onderrijk Dikarya. Een vergelijkbare levenscyclus heeft zich onafhankelijk bij de roodwieren ontwikkeld.
Uit het bevruchte ascogonium vormen zich nu miljoenen 'dinucleate' schimmeldraden (hyfen), die in iedere cel twee celkernen hebben. Ze worden ook 'fertiel' of 'ascogeen' genoemd en worden gevoed door gewone schimmeldraden met één celkern ('uni-' of 'mononucleaat'), die als 'steriel' aangeduid worden. Het weefsel van fertiele en steriele schimmeldraden vormt nu het ascocarp van de schimmel, dat in vele gevallen met het oog zichtbaar is.

Op het binnenste oppervlak van het ascocarp bevindt zich het hymenium, een vruchtlaag waarin sporenzakjes ontstaan. Daartoe vormt een 'ascogene hyfe' aan het uiteinde een haak, tegen de groeirichting van de schimmeldraad in. De cellen in deze haak bevatten twee celkernen en ze delen zich dusdanig dat de kernspoelen van hun mitosespoelen parallel verlopen waardoor aan het einde van de haak twee dochterkernen komen te liggen die genetisch van elkaar verschillen. Een daarvan bevindt zich aan het uiteinde van de haak. De andere dochterkern is gepositioneerd aan de basis van de hyfecel. Vervolgens worden twee tussenwanden aangemaakt, die het uiteinde van de schimmeldraad in drie delen verdelen: de haak met één kern, de daaronder gelegen basis van de schimmeldraad, eveneens met één kern, en een nieuw uiteinde van de schimmeldraad met twee celkernen. Bij de laatste vindt bij de juiste positionering in de vruchtlaag uiteindelijk de kernversmelting plaats, waardoor de diploïde zygote ontstaat. Deze verlengt zich nu tot een langwerpig band- of cilindervormig kapsel, het eigenlijke sporenzakje.
Hier vindt de deling (meiose) plaats, waardoor het aantal kernen naar vier stijgt en tegelijkertijd de haploïde toestand hersteld wordt. Bijna altijd volgt een verdere kerndeling (mitose) zodat uiteindelijk acht dochterkernen in het sporenzakje komen te liggen. Deze worden omhuld met wat celplasma van eigen membranen en meestal ook met een vaste celwand. Zo vormen zich ten slotte de eigenlijke voortplantingscellen, de ascosporen. Deze liggen in de sporenzakjes als erwten in een peul en worden op een passend moment losgelaten.
Ascosporen kunnen in principe niet zelfstandig bewegen, waardoor ze voor verspreiding van externe mechanismen afhankelijk zijn. Vele sporen worden door middel van de wind verspreid, bij andere breken de rijpe sporenzakjes bij contact met water open. Enkele soorten hebben "sporenkanonnen", waarmee ze hun sporen tot 30 cm ver weg kunnen schieten. Schimmels uit het geslacht Ascobolus beschikken over een lenzensysteem waarmee de 'loop', het sporenzakje, van het sporenkanon[* 6] gericht wordt op het invallende licht, zodat de sporen in een turbulente luchtzone geschoten worden.
Zodra de sporen op een geschikt substraat terechtkomen kiemen ze, vormen nieuwe schimmeldraden en starten zo de levenscyclus van voren af aan.
Samengevat bestaat de levenscyclus uit drie generaties (trigenetische cyclus):
- haploïde schimmeldraden afkomstig van ascosporen
- de dikaryofase, waar - na versmelting van twee passende schimmeldraden - elke cel twee kernen heeft
- een (kortdurende) diploïde sporofytfase, die beëindigd wordt door de vorming van haploïde ascosporen bij de meiose
Classificatie van de sporenzakjes[bewerken | brontekst bewerken]
Bij de classificatie van de zakjeszwammen speelt ook de vorm van het sporenzakje een rol. Er zijn vier typen te onderscheiden, afhankelijk van het aantal wanden van het zakje en het verspreidingsmechanisme van de sporen:
- Een 'unitunicaat operculaat' sporenzakje is enkelwandig en heeft een ingebouwde dop, het 'operculum'. Daarin zit een zwakke plek waar het zakje bij rijping van de sporen openbreekt zodat ze losgelaten worden. Unitunicaat-operculate sporenzakjes komen alleen voor bij ascocarpen van het apothecium-type, bijvoorbeeld bij morieltjes.

- Een 'unitunicaat inoperculaat' sporenzakje is eveneens enkelwandig, maar bezit geen operculum. In plaats daarvan is er een elastische ring die als ventiel functioneert. Bij rijping ontspant deze zich kortdurend en laat zo de sporen ontsnappen. Dit type vindt men zowel bij apotheciën als bij peritheciën. Een voorbeeld is de goudgele zwameter (Hypomyces chrysospermus).
- Een 'bitunicaat' sporenzakje wordt omgeven door een dubbele wand. Deze bestaat uit een dunne, broze buitenwand en een dikke, elastische binnenwand. Zodra de sporen rijp zijn splijt de buitenwand open zodat de binnenwand water kan opnemen. Als gevolg hiervan begint deze samen met de sporen uit te zetten totdat de wand boven het ascocarp uit komt, zodat de sporen ongehinderd door de vrije luchtstroom meegenomen kunnen worden. Bitunicate sporenzakjes treden uitsluitend op bij pseudotheciën, zoals appelschurft en Guignardia aesculi.
- Sporenzakjes die niet tot een van voorgaande typen horen vallen onder de verzamelaanduiding 'prototunicate' sporenzakjes. Veelal zijn deze kogelvormig en ze hebben doorgaans geen actief verspreidingsmechanisme. De wand van het sporenzakje lost simpelweg op, zodat de sporen eruit kunnen stromen, dan wel wordt deze opengebroken door externe invloeden, zoals dieren. Protunicate sporenzakjes komen zowel voor bij peritheciën als bij cleistotheciën, zoals bij de iepziekte en de truffels. Waarschijnlijk hebben vele prototunicate sporenzakjes zich onafhankelijk van elkaar uit unitunicate sporenzakjes ontwikkeld.
Voortplanting bij de Taphrinales[bewerken | brontekst bewerken]
Taphrinales onderscheiden zich van andere zakjeszwammen door hun tweekernige schimmeldraadcellen. Ze hebben ook een karakteristieke manier van voortplanting. Onder de cuticula van het blad waarin ze leven vormt zich een vlechtwerk van schimmeldraden, dat een proascus wordt genoemd. De twee haploïde kernen in een schimmeldraadcel versmelten tot een diploïde cel die de cuticula doet openbarsten. De diploïde cel wordt naar buiten gedrukt en vormt vervolgens een scheidingswand, waardoor twee cellen ontstaan. De onderste cel is de voetcel en de bovenste cel het sporenzakje. In het sporenzakje vindt de meiose plaats, gevolgd door een mitotische deling. Dat resulteert in acht haploïde ascosporen. Deze maken door knopvorming net zo lang secundaire sporen (conidiën) totdat ze op de gastheer komen. Dan vormt de secundaire spore een kiembuis, die tussen de epidermiscellen van het blad groeit. Vervolgens groeien door het gehele blad intercellulair schimmeldraden.
Betekenis voor de mensheid[bewerken | brontekst bewerken]
Zakjeszwammen zijn voor de mensheid van grote betekenis. Ze hebben veel effecten op de leefomgeving van de mens, zowel nuttige als schadelijke.
Schadelijke effecten[bewerken | brontekst bewerken]
Talrijke ziektes bij mensen, huisdieren, vee en cultuurgewassen worden veroorzaakt door zakjeszwammen. Ook kunnen ze celluloserijke bouwmaterialen en textiele stoffen aantasten.
Plantenziektes[bewerken | brontekst bewerken]


Een van de belangrijkste schadelijke effecten van zakjeszwammen is de verspreiding van talrijke plantenziektes. De grootschalige iepensterfte die sinds de jaren zeventig van de vorige eeuw Noord-Amerika en Europa teistert, wordt bijvoorbeeld veroorzaakt door de nauw verwante soorten Ophiostoma ulmi en Ophiostoma novo-ulmi.
Kastanjekanker bij tamme kastanjes wordt veroorzaakt door de uit Azië stammende schimmelsoort Cryphonectria parasitica. Een ziekte van maïsplanten die in het bijzonder in Noord-Amerika heerst wordt veroorzaakt door Cochliobolus heterostrophus. Taphrina deformans verspreidt de perzikkrulziekte. Erysiphe necator is voor de aantasting van wijnstokken met echte meeldauw verantwoordelijk, terwijl ascomyceten van het geslacht Monilia in het bijzonder steenvruchten aantasten. Daarnaast worden ook de bloesems van perzik en zure kers geïnfecteerd. Schurft (Venturia inaequalis) is een van de belangrijkste plantenziekten op appel.
Blauwgroene, rode en bruine schimmels kunnen levensmiddelen bederven, Penicillium digitatum en Penicillium italicum bijvoorbeeld sinaasappels.
Dierziektes[bewerken | brontekst bewerken]
Katten, honden, paarden en koeien lopen vaak huidinfecties op die veroorzaakt worden door zakjeszwammen die behoren tot de geslachten Microsporum en Trichophyton. Schimmelinfecties kunnen er bij runderen en paarden, onder andere door Aspergillus fumigatus, toe leiden dat de ongeboren vrucht verworpen wordt (spontane abortus). Graan dat geïnfecteerd is met Fusarium graminearum bevat mycotoxinen als deoxynivalenol (DON), dat bij opname kan leiden tot huid- en slijmvliesletsel bij varkens.
Ziekte bij de mens[bewerken | brontekst bewerken]
Van direct belang voor de mensheid is de moederkoornschimmel (Claviceps purpurea), die rogge en gewone tarwe aantast en zeer giftige en mogelijk zelfs kankerverwekkende alkaloïden vormt, zoals ergotamine. Deze kunnen bij opname onder andere darmkrampen en zware hallucinaties veroorzaken. Zeer kankerverwekkend en leverbeschadigend is de stof aflatoxine, die wordt afgescheiden door Aspergillus flavus, een ascomyceet die onder andere op pinda's groeit. In vergelijking daarmee is de gistschimmel Candida albicans, die menselijke slijmvliezen kan aantasten en daarbij candidiasis veroorzaakt, redelijk onschadelijk.
Ook de voor infecties verantwoordelijke huidschimmels van het geslacht Epidermophyton zijn voor mensen met een gezond immuunsysteem relatief ongevaarlijk. Is het immuunsysteem echter verstoord, dan kunnen de schimmels tot levensgevaarlijke ziektes leiden; infectie met Pneumocystis jiroveci kan bijvoorbeeld zware longaandoeningen tot gevolg hebben. Trichophyton rubrum is een schimmel die bekendstaat als veroorzaker van zwemmerseczeem en onychomycose.
Daarnaast zijn er schimmels die sporen produceren die bij het inademen een schimmelinfectie kunnen veroorzaken. Na infectie komt de schimmel dan voor in gistvorm. Voorbeelden hiervan zijn histoplasmose, dat veroorzaakt wordt door de schimmel Histoplasma capsulatum, en blastomycose, veroorzaakt door de schimmel Blastomyces dermatitidis.
Aantasting materialen[bewerken | brontekst bewerken]
Stachybotrys chartarum komt veel voor in door vocht en wateroverlast aangetaste gebouwen en zorgt in het bijzonder in de tropen voor grote problemen.
Natuurlijke textiele stoffen kunnen worden aangetast door verschillende schimmels, waaronder Aspergillus flavus, Aspergillus fumigatus, Aureobasidium pullulans, Aspergillus niger en Alternaria alternata.[20] Waarvan Aureobasidium pullulans en Aspergillus niger het meest voorkomen.[21] De aantasting is onder meer zichtbaar als het weer.
Nuttige effecten[bewerken | brontekst bewerken]
Zakjeszwammen kunnen ook heel nuttige effecten sorteren. Zo spelen ze een belangrijke rol bij de productie van medicijnen en de bereiding van levensmiddelen. Sommige dienen zelf als voedsel en weer andere vormen een bestrijdingsmiddel tegen insecten.
Medicijnen[bewerken | brontekst bewerken]
Het bekendste voorbeeld van een zakjeszwam die een medicijn maakt is de soort Penicillium chrysogenum. Deze schimmel produceert een antibioticum dat vermoedelijk dient om concurrerende bacteriën te bestrijden, maar dat als penicilline in de 20e eeuw een revolutie in de behandeling van bacteriële infectieziekten veroorzaakt heeft.
Ook van grote betekenis is Tolypocladium inflatum, een ascomyceet die met het geneesmiddel ciclosporine een van de effectiefste en mooiste middelen voor immuunsysteemonderdrukking mogelijk maakt. Behalve als middel om bij orgaantransplantatie afstoting van lichaamsvreemde weefsels te voorkomen, wordt ciclosporine ook gebruikt voor behandeling van auto-immuunziektes zoals multiple sclerose. Hier is echter het risico van zich op langere termijn ontwikkelende ernstige bijwerkingen aanwezig.
Daarnaast kunnen vele ascomyceten verhoudingsgewijs eenvoudig genetisch gemanipuleerd worden. Ze produceren dan belangrijke eiwitten als insuline, menselijke groeifactoren en tPA, een stof die ingezet wordt voor het oplossen van bloedproppen. De broodschimmel Neurospora crassa, waarvan het genoom ondertussen volledig in kaart is gebracht, is een belangrijk modelorganisme in de biologie.
.jpg)

Levensmiddelen[bewerken | brontekst bewerken]
Zakjeszwammen worden veelvuldig ingezet bij de productie van levensmiddelen. Belangrijk is bakkersgist dat vaak wordt gebruikt bij het bakken van onder andere brood. Bij voldoende zuurstofaanbod kan het afgegeven koolstofdioxide benut worden om het deeg te laten rijzen. Het gist wordt ook benut bij de bereiding van bier en wijn, waarbij de suikers zoals glucose, fructose of sacharose vergist worden tot alcohol.
Enzymen van Penicillium camemberti spelen een rol bij de productie van de kaassoorten Camembert en Brie, en enzymen van Penicillium roqueforti bij de vervaardiging van Gorgonzola, Roquefort en Stilton. Aspergillus oryzae wordt in het bijzonder in Azië ingezet voor vergisting van een brij uit geweekte sojabonen; door de vergisting ontstaat de in vele oosterse gerechten gebruikte sojasaus. Aspergillus niger wordt gebruikt voor de industriële productie van citroenzuur en gluconzuur en van vele enzymen, zoals pectinase, amylase en protease.
Zakjeszwammen kunnen gebruikt worden bij de biologische bestrijding van insecten die schadelijk zijn bij de teelt van gewassen. Zo is Beauveria bassiana een ascomyceet, die over de gehele wereld in de bodem voorkomt. De schimmel ontwikkelt zich in het lichaam van de geleedpotige en veroorzaakt zijn dood. De schimmel is niet schadelijk voor mens of milieu.
Ten slotte zijn er ook ascomyceten met eetbare paddenstoelen. Hoewel de steeltjeszwammen wat dit betreft getalsmatig belangrijker zijn, behoren morieltjes en truffels, de twee culinair meest gewaardeerde paddenstoelen, tot de zakjeszwammen.
Bedreiging met uitsterven[bewerken | brontekst bewerken]
Anders dan bij zoogdieren en vogels is er in de regel te weinig over de populaties van de verschillende zakjeszwammen bekend om zinnige uitspraken te kunnen doen over de mate waarin ze bedreigd worden met uitsterven. Dit geldt in het bijzonder voor de talrijke wortelschimmels, die belangrijke ecologische taken hebben en vaak alleen microscopisch waargenomen kunnen worden. Echter ook bij grotere schimmels is de beschikbare informatie zeer spaarzaam. Pas in 2003 zijn op de rode lijsten ook schimmels gezet. Sinds 2016 staan er negen ascomyceten-soorten op de Rode Lijst van de IUCN met bedreigde schimmels.[22]
In België bestaan vooralsnog geen rode lijsten voor ascomyceten. Nederland kent er twee, een rode lijst van korstmossen en een rode lijst van paddenstoelen. Op de lijst van korstmossen staan 323 soorten, een aanzienlijk deel van de in totaal ongeveer 750 soorten die in Nederland voorkomen. Om de verspreiding van kortmossen te meten bestaat op de zandgronden sinds 1988 een meetnet waarbij iedere vijf jaar 30 000 bomen bemonsterd worden. Korstmossen zijn erg gevoelig voor luchtverontreiniging en daardoor bruikbaar als instrument om de verontreiniging te meten.
Evolutie[bewerken | brontekst bewerken]
In het Devoon, ongeveer 400 Ma geleden, zijn de stammen Ascomycota en Basidiomycota uit een gemeenschappelijke voorouder ontstaan.[23][24] Mogelijk zijn de Ascomycota 50 Ma ouder. In fossielen afkomstig uit het vroege Siluur in Virginia en die uit het late Siluur in Zweden zijn namelijk op fialiden lijkende structuren gevonden.[25]
Uit een fylogenetisch onderzoek dat verricht werd met behulp van zes genen bij 420 schimmelsoorten, blijkt dat zakjeszwammen (Ascomycota) en steeltjeszwammen (Basidiomycota) zich ontwikkeld hebben uit saprofyten. Bovendien bleek dat korstmossen diverse malen in de evolutie zijn ontstaan (licheenvorming).[26]

In de Rhynie Chert, een bekende vindplaats van plantenfossielen in het Schotse Aberdeenshire, werden in doorsneden onder de epidermis van de verticale en horizontale stengeldelen (telomen) van de vroegere landplant Asteroxylon mackiei sporenzakjes, ascosporen en peritheciën geïdentificeerd, die zich eenduidig met ascomyceten in verbinding laten brengen. Het gaat hier om de schimmelsoort Paleopyrenomycites devonicus. Daarmee staat vast dat wisselwerkingen tussen schimmel en plant al zeer vroeg zijn ontstaan; er wordt zelfs gespeculeerd dat de enge symbiotische relatie tussen planten en schimmels een voorwaarde kan zijn geweest voor het koloniseren van het land. Interessant is dat op de vindplaats bij Rhynie ook de oudste korstmossen zijn gevonden.
Een fossiel uit het Eoceen dat gevonden werd in de Messelgroeve in Noord-Duitsland toont een mier, die zich heeft vastgebeten in een blad. Dit duidt erop dat de mier geïnfecteerd was met een schimmel, vergelijkbaar met het nu nog voorkomende parasitisme bij mieren.[27]
Systematiek[bewerken | brontekst bewerken]

De groep Ascomyceten omvat alle nakomelingen van een gemeenschappelijke voorouder en vormt zo een monofyletisch taxon. Vooral op grond van de synapomorfie van het sporenzakje, dat alleen bij zakjeszwammen optreedt, kan dit als vrijwel vaststaand beschouwd worden. Bij schimmels zonder geslachtelijk stadium is een indeling bij de ascomyceten veelal mogelijk op grond van eenduidige moleculairbiologische of ultrastructurele gegevens.
De indeling van zakjeszwammen met steeltjeszwammen in een hogere groep, die men Dikarya noemt, is gebaseerd op de regelmatige septering van de hyfen door tussenwanden met centrale perforatie, die kenmerkend is voor beide. De naam "dikarya" refereert aan de dikaryon-fase in beider levenscyclus, alsook aan de daaropvolgende versmelting van celplasma's. Bovendien kan bij beide groepen fusie optreden van steriele hyfen (anastomose). Bij ascomyceten ontwikkelen zich, kort voor de sporenzakjesvorming, aan het uiteinde van de ascogene schimmeldraden U-vormige haken, terwijl bij basidiomyceten gespen aangetroffen worden. Deze twee kenmerken worden als homologiën gezien, als structuren die vermoedelijk afkomstig zijn van een gemeenschappelijke voorloperstructuur.
De ascomyceten vertegenwoordigen 75 procent van alle echte schimmelsoorten. Er werden tot nog toe meer dan 64 000 soorten beschreven, waarvan ruim 40 procent bestaat uit korstmossen. In bepaalde, weinig onderzochte ecosystemen zoals in tropische gebieden, is de schimmelflora nog onvoldoende bekend. Waarschijnlijk is het werkelijke aantal soorten een veelvoud van het getal dat nu gehanteerd wordt.
Een groot probleem bij de taxonomische indeling van de schimmels (Fungi) is de te volgen benadering. De indeling verandert voortdurend door een toegenomen interesse, betere waarnemingstechnieken, betere apparatuur, nieuw ontdekte soorten en een beter begrip van de relaties tussen schimmels.
Klassiek werden schimmels vooral geclassificeerd op basis van hun morfologie en levenscyclus, de zogenaamde verticale classificatie. Kenmerken als vorm, afmeting en kleur spelen daarbij een belangrijke rol, evenals de aan- of afwezigheid van bepaalde kenmerken. Het gebruik van kenmerken die meermaals in de geschiedenis ontstaan zijn zorgde voor een dubieuze, soms zeer kunstmatige indeling.
Recent worden ook scheikundige en genetische technieken gebruikt die de schimmels classificeren op basis van hun samenstelling. Deze horizontale classificatie geeft directere informatie over de onderlinge soortverwantschappen en wordt ook bij ascomyceten toegepast. Door de toegenomen kennis van de ascomyceten wordt de kunstmatige indeling geleidelijk verlaten en kiest men voor een indeling van zakjeszwammen in natuurlijke taxa. Een veel gebruikte indeling in klassen wordt gegeven door de 'Index Fungorum'.[* 7]
Taxonomie[bewerken | brontekst bewerken]
Holt en Iudica gaven in 2016 de volgende bijgewerkte indeling:[2]
Oudere indelingen[bewerken | brontekst bewerken]
Vroeger werd gebruikgemaakt van een indeling op basis van het ascocarp van de schimmel:
- Euascomyceten: de ascomyceten die wel echte ascocarpen ontwikkelen.
Dit is de grootste groep, die ook de farmaceutisch meest belangrijke schimmels bevat (zoals Penicillium chrysogenum), alsook de truffels en morieljes. - Hemiascomyceten: de groep ascomyceten die geen echte ascocarpen ontwikkelen.
Ascocarpen ontwikkelen zich direct uit de ascogene hyfen, zonder het tussenstadium van gespecialiseerde cellen. Men maakt nog een verder onderscheid tussen de:- Discomyceten: vorming van apothecium
- Pyromyceten: vorming van perithecium
- Loculoascomyceten: vorming van pseudothecium
- Plectomyceten: vorming van cleistothecium
- Archaeoascomyceten: deze hebben geen ascocarpen; ze vormen een relatief kleine groep, die traditioneel tot de hemiascomyceten gerekend werd.
De Archaeoascomyceten hebben zich wellicht vroeg los van de andere ascomyceten ontwikkeld als een afzonderlijke natuurlijke groep, maar deze stelling is zeer omstreden.
In onderstaande, oudere indelingen zijn de klassen niet gegroepeerd in onderstammen (subfyla). De Pezizomycotina hebben daar nog de naam Ascomycetes.
Onderzoek[bewerken | brontekst bewerken]
De systematiek van zakjeszwammen is nog altijd een belangrijk onderzoeksgebied. De plaatsing van de ascomyceten als zustergroep van de basidiomyceten is weliswaar relatief onomstreden, vragen naar de verdere indeling zijn echter nog steeds actueel. In het bijzonder de verhouding van de archaeoascomyceten tot de rest van ascomyceten is nog niet opgehelderd; ook de gedetailleerde classificatie binnen de ascomycetenordes is nog in beweging.
Een ander vruchtbaar onderzoeksgebied is de bestudering van de afstamming van de ascomyceten aan de hand van fossiele sporen, asci of hele ascocarpen. In het bijzonder de associatie van ascomyceten met de wortels van de eerste landplanten is bij verre nog niet opgeklaard; nieuwe fossielenvondsten zijn dringend nodig om de landkolonisatie van planten en schimmels beter te begrijpen.
De rol van zakjeszwammen in ecosystemen is een ander onderwerp van studie in de schimmelkunde. In het bijzonder de endofyten en hun uitwerking op planten zijn beperkt onderzocht. Terwijl de lichenologie (korstmossenkunde) traditioneel een zelfstandige wetenschap is, valt het onderzoek naar wortelschimmelsymbioses en endofytische schimmels in het werkgebied van mycologen.
Verscheidene schimmels dienen als modelorganismen in de genetica en moleculaire biologie. Onder andere op Neurospora crassa[31] en het bakkers- of biergist (Saccharomyces cerevisiae)[32] wordt onderzoek gedaan naar talrijke celbiologische fenomenen, die van elementaire betekenis zijn voor de kennis van alle organismen en de daaraan ten grond liggende levensprocessen.
De vraag van de soortbescherming komt steeds meer op de voorgrond: De bedreigingsstatus van een soort kan alleen goed beoordeeld worden indien men degelijke gegevens over de bestandsontwikkeling heeft - deze bestaan tegenwoordig vaak alleen in uitzonderingsgevallen, zelfs op de gematigde breedtes.
Anno 2016 is nog slechts een gering deel van alle ascomyceten beschreven. De beschrijving van nieuwe soorten, in het bijzonder uit de tropen, zal de mycologie nog lang bezighouden.
Noten
Literatuur
Referenties en verwijzingen
|
| Dit artikel is op 2 augustus 2016 in deze versie opgenomen in de etalage. |